НОВЫЕ ЗВЕНЬЯ В ПАТОГЕНЕЗЕ ХРОНИЧЕСКИХ ИНФЕКЦИЙ
("Врач". 1997.- N 7.- С. 40-41)
Новосибирская государственная медицинская академия, ЦНИЛ
Патофизиологические
механизмы, определяющие развитие и течение многих хронических инфекций
(туберкулез, лепра, бруцеллез), до конца не раскрыты. Самым
загадочным из них и неизученным является механизм торможения
в макрофагах процесса слияния фагосом, содержащих бактерии, с лизосомами.
Многочисленные попытки отечественных и зарубежных исследователей
объяснить этот механизм не
увенчались успехом. Например, в монографиях
А.Н.Маянского и Д.Н.Маянского [9], А.Г.Булычева [4], Д.Н.Маянского
[10] приведено много литературных и собственных данных о том, что
при захвате макрофагами микобактерий (МБ) почему-то
не происходит фагосомно-лизосомного слияния (ФЛС). Они спокойно живут
в макрофагах и размножаются. До сих пор не удалось создать
эффективных препара-
тов, стимулирующих ФЛС. Такое положение дел, сложившееся в учении о
воспалении, обусловлено отсутствием адекватных концептуальных и патофизиологических
моделей взаимодействия бактерий с фагоцитами на уровне очага воспаления.
Так как фагоцитоз изучали в основном на культурах клеток в искусственных
условиях, то эти искусственные патофизиологические модели породили
подобные им концептуаль-
ные модели. Основной целью этих работ было выяснение молекулярных механизмов
фагоцитоза. За короткий промежуток времени исследователям удалось
разобраться в этих механизмах, однако односторонняя увлеченность молекулярной
биологией отвлекла их от изучения тканевых и межклеточных взаимодействий.
Между клеточно-тканевым и молекулярным уровнем
исследования образовалась информационная пропасть, так
как из-за увлеченностью молекуляными механизмами были не до
конца изучены клеточные и тканевые особенности течения острого и хронического
воспаления. В патогенезе хронических инфекций было упущено
несколько важных звеньев, определяющих механизм невосприимчивости
макрофагов к МБ.
По мнению автора статьи,
"невосприимчивость" макрофагов к МБ, возникает после
взаимодействия бактерий с полиморфноядерными лейкоцитами (ПЯЛ) [1,2,3].
Как известно [9,10,11], фагоцитоз бактерий ПЯЛ происходит в острой
стадии заболевания и в периоды обострения.
В эти две ключевые стадии формируются основные патологические замкнутые
круги, способствующие паразитированию бакте-
рий в макрофагах (эндоцитобиоз).
Выяснено [9,11],
что ПЯЛ фагоцитируют МБ, попавших во внутреннюю среду организма человека,
но уничтожить их они не в состоянии. Несмотря на то, что ПЯЛ принимают
активное участие в одном из ключевых звеньев патогенеза микобактериозов,
клиницисты и ученые практически не обращают на них внимания. Например,
Г.В.Ковалевский [6] вводил внутрибрюшинно крысам
БЦЖ и изучал реакцию лимфатических узлов и селезенки. В результате, он
обнаружил через
1-2 суток после введения БЦЖ в синусах регионарных лимфатических
узлов лейкофагию. Описав ее, он, однако, не делает никаких выводов
и попыток объяснить данный феномен. В связи с этим, возникает вопрос: какова
роль ПЯЛ в патогенезе микобактериозов? Если мы
рассмотрим банальное остро протекающее инфекционное заболевание,
то решающую роль ПЯЛ в данном случае никто не отрицает. Но когда
речь заходит о микобактериозах, то роль ПЯЛ принижается. Уже тот
факт, что они не могут справиться с МБ, заставляет признать
их ключевым звеном в патогенезе этих заболеваний. Данное
ключевое звено играет огромную роль в начальной, острой стадии микобактериозов.
Реально же врач-клиницист сталкивается с подострой и хронической стадией.
Невозможность, в подавляющем большинстве случаев, зафиксировать у больного
цито-морфологическую картину начальной стадии инфицирования и явилось причиной
недооценки роли ПЯЛ в патогенезе микобактериозов. Исследователями был упущен
важный механизм - процесс конвеерной упаковки бактерий в мембранные
оболочки. В упаковке бактерий участвуют два типа фагоцитов:
ПЯЛ и макрофаги; причем ПЯЛ начинают ее, а макрофаги продолжают.
Опишем
процесс конвеерной упаковки бактерий в мембранные оболочки (см. рис.1).
На рисунке светлые стрелки указывают на последовательность формирования
в макрофаге внутриклеточной трехмембранной упаковки МБ, а темные
- на образование одно- и двухмембранных упаковок. Как видно
из рисунка, процесс взаимодействия МБ с фагоцитами начинается с фагоцитоза
микобактерии (1) ПЯЛ с образованием в нем внутриклеточной одномембранной
формы упаковки МБ (3). Это типичная фагосома с МБ. В дальнейшем ПЯЛ
распадается, с выбросом в межклеточную среду неупакованных (1) и упакованных
(2) МБ или его целиком фагоцитирует макрофаг и в
нем формируется внутриклеточная трехмембранная форма
упаковки МБ (5). В данном случае МБ окружена тремя замкнутыми мембранными
оболочками, две из которых являются оболочками фагосом (наружная
и внутренняя), а третья (средняя) - наружной клеточной мембраной ПЯЛ.
Остатки содержимого разрушенного ПЯЛ также фагоцитирует макрофаг,
и в нем формируются внутриклеточная одно- и двухмембранная
упаковки МБ (3,4). Первая из них является типичной фагосомой,
а вторая - фагосомой с двумя фагосомными мембранами (внутренняя - ПЯЛ,
наружная - макрофага).
Все это позволяет
выделить в процессе взаимодействия бактерий с фагоцитами несколько
патофизиологических звеньев и кратко охарактеризовать их роль в патогенезе
микобактериозов:
1. Микобактерию фагоцитирует ПЯЛ,
и она находится в нем в одномембранной упаковке (фагосоме). Затем,
макрофаг фагоцитирует этот ПЯЛ, и в макрофаге МБ уже находится в трехмембранной
упаковке. Происходит ФЛС. Лизосомальные ферменты разрушают
внутренние структуры этой фагосомы, и МБ получает полный комплекс субстратов
для своей жизнедеятельности.
2. Микобактерию фагоцитирует ПЯЛ, разрушается,
и в межклеточном пространстве оказываются его фрагменты с фагосомами,
содержащими МБ. Эти фагосомы, в свою очередь, фагоцитирует
макрофаг, и МБ оказывается в двухмембранной внутриклеточной упаковке.
Происходит ФЛС. остатки ПЯЛ разрушаются, а МБ, как и
в первом случае, получает полный комплекс субстратов
для своей жизнедеятельности.
3. Микобактерию фагоцитирует макрофаг
и, не справившись, разрушается. Фрагменты его
и фагосому с МБ фагоцитирует новый макрофаг. Формируется двухмембранная
упаковка МБ. Происходит ФЛС. Формируется одномембранная упаковка
с комплексом питательных субстратов для МБ.
4. Микобактерию фагоцитирует макрофаг и разрушает.
В дальнейшем
необходимо выяснить роль каждого из вышеперечисленных вариантов патогенеза.
Особо следует отметить тот факт, что бактерии могут находиться в межклеточном
пространстве свободно или в мембранной упаковке (в фагосоме) [12].
Это приводит к формированию двух основных звеньев патогенеза,
так как в первом случае макрофаг реагирует на МБ (чужеродный объект),
а во втором - на фагосому (одна из структур организма), в которой
антигенные детерминанты МБ скрыты от иммунной системы
(эффект "Троянского коня").
В заключении
следует отметить то, что человек и окружающая его среда едины,
и частным отображением этого единства является симбиоз человека с
микробами [8]. Согласно И.В.Давыдовскому [5], существует два вида
симбиоза - физиологический и патологический (когда сапрофит
становится паразитом). В патогенезе хронических
инфекций всегда присутствует стадия симбиоза. Например, известно, что
почти все люди инфицированы микобактериями туберкулеза.
У большинства из них нет выраженного клинического проявления заболевания,
т.к. у них развился фзиологический симбиоз. На это указывает наличие в
организме внешне здоровых людей L-форм микобактерий [7].
Кроме этого, фтизиатры широко используют симбиоз в своей повседневной
деятельности (вакцинация БЦЖ). Поэтому одной
из главных задач при лечении микобактериозов является не
только достижение состояния абациллирования, но и перевод патологического
симбиоза (острого и хронического воспаления) в физиологический. А
главной задачей теоретической медицины - расшифровка механизмов становления
симбиоза микобактерий с макроорганизмом. При этом ключевым звеном
становления симбиоза, как указывалось выше, является процесс
конвеерной упаковки бактерий в мембранные оболочки.
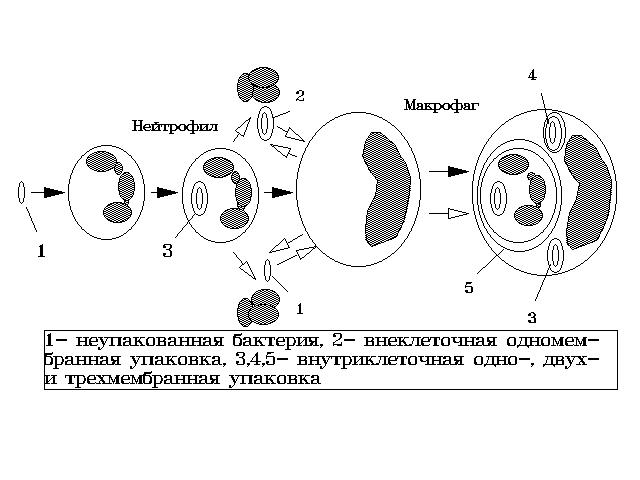
Рис.1. Схема конвейерной упаковки бактерий
в мембранные оболочки.
СПИСОК ЛИТЕРАТУРЫ
1. Белкин А.Д. Микобактерии в мембранной
упаковке//Научная сессия посвященная 100-летию Новосибирска,
25 - 28 января 1994 г.: Тез. докл.
Новосибирск, 1994. - С. 177.
2. Белкин А.Д. Патофизиологические
механизмы формирования эндоцитосимбиоза/Актуальные вопросы
патофизиологии лимфатической
системы. Новосибирск, 1995.- С. 74 - 77.
3. Белкин А.Д. Один из возможных механизмов
взаимодействия микобактерий с фагоцитами/ 70 лет противотуберкулезной службе
Новосибирской области. 50 лет Новосибирскому НИИ туберкулеза:
сб. науч. тр. Юбилейной науч.-практич. конф., Новосибирск, июнь 1995. -
Новосибирск. - 1995.- С. 299 - 301.
4. Булычев А.Г. Сегрегационная функция
клетки.-Л.: Наука, 1991.- 112 с.
5. Давыдовский И.В. Учение
об инфекции.- М.: Медицина, 1956.- 107 с.
6. Ковалевский Г.В. Очерки иммунологии
на модели микобактериальной сенсибилизации резистентных к туберкулезу животных.
Новосибирск, изд-во Наука, Сиб. от-ние, 1976.-226 с.
7. L-формы микобактерий туберкулеза/ Под ред.
З.Н.Кочемасо-
вой.-М.: Медицина, 1980.-174 с.
8. Маргелис Л. Роль симбиоза в
эволюции клетки: Пер. с англ.-М.: Мир.-1983.-352 с.
9. Маянский А.Н., Маянский Д.Н. Очерки о нейтрофиле
и макрофаге.- Новосибирск: Наука, Сиб. отд-ние, 1989. - 344 с.
10. Маянский Д.Н. Хроническое
воспаление.-М.: Медицина,1991.-272 с.
11. Руководство по туберкулезу/Под ред.
С.М.Княжецкого.-Л.: Медицина, Ленинградское отд-ние, 1972. - 623 с.
12 . Ультраструктурная патология инфекционных болезней/Б.А.Ерман,
Л.Г.Тулакина, В.П.Устьянцев и др.- Новосибирск: Наука. Сиб. отд-ние, 1992.
- 92 с.