На правах рукописи
БЕЛКИН
Анатолий Дмитриевич
СТРУКТУРНО-ФУНКЦИОНАЛЬНЫЕ ИЗМЕНЕНИЯ В ОРГАНИЗМЕ
ПРИ ВОЗДЕЙСТВИИ ТЕХНОГЕННЫХ ВРАЩАЮЩИХСЯ
И ПЕРЕМЕННЫХ ЭЛЕКТРИЧЕСКИХ ПОЛЕЙ
И МЕХАНИЗМЫ ИХ ВОЗНИКНОВЕНИЯ
03.00.16 - экология
Автореферат
диссертации на соискание ученой степени
доктора биологических наук
Новосибирск - 1999
Работа выполнена в Новосибирском медицинском институте
МЗ РФ.
Научный консультант:
доктор медицинских наук, профессор А.Н.Мосолов
Официальные оппоненты:
доктор медицинских наук, профессор В.Ю.Куликов
Заслуженный работник сельского хозяйств РФ,
доктор биологических наук, профессор А.Г.Незавитин
Заслуженный деятель науки РФ,
доктор биологических наук, профессор Н.О.Сухова
Ведущая организация:
Новосибирский государственный педагогический университет.
Защита состоится "___" ___________1999 г. в ___час. на заседа-
нии Диссертационного совета Д 120.32.02 в Новосибирском
государственном аграрном университете (630039; Новосибирск,
ул. Добролюбова, 160)
С диссертацией можно ознакомиться в библиотеке Новосибирского
государственного аграрного университета
Автореферат диссертации разослан " "
1999 г.
Ученый секретарь
Диссертационного совета,
кандидат биологических наук
Кочнев Н.Н.
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актульность проблемы. Всего за несколько десятков лет, незаметно,
появился новый урбоэкологический фактор - вращающееся электрическое поле
(ВЭП). Оно возникло, в окружающей среде, благодаря использованию в промышленности
и быту переменного трехфазного электрического тока и переменных электромагнитных
полей (ЭМП). Еще 100 лет назад в биосфере преобладали природные статические
и переменные электрические поля (ЭП). И вот за короткий исторический период
произошло резкое изменение ее электромагнитного фона. В настоящее время
большинство жителей городов круглосуточно находятся во ВЭП. Особо подвержены
воздействию этого вида ЭП рабочие, обслуживающие линии электропередачи
(ЛЭП) и высоковольтные установки (Пучков Г.Г. и др., 1986).
До последнего времени факт появления в биосфере ВЭП не принимался во
внимание. Поэтому совершенно не изучены механизмы взаимодействия нового
урбоэкологического фактора со здоровым и больным организмом.
Особенно актуальным, с точки зрения эндоэкологии, нормальной и патологической
физиологии, является раскрытие роли эндогенного ВЭП в жизнедеятельности
организма. Его создает электродвижущая сила сердечной мышцы (Тарковский
М.Б., 1964; Орлов В.Н., 1983). До настоящего времени физиологи не обращали
внимания на форму ЭП, генерируемого сердцем, так же как экологи не обращали
внимания на форму ЭП ЛЭП. Как осуществляется процесс взаимодействия экзогенного
ВЭП с эндогенным, и каковы последствия этого взаимодействия, никто не знает.
В отечественной и зарубежной научной литературе имеются многочисленные
данные о реакциях биосистем на воздействие внешних ЭП и очень мало работ
посвящено изучению механизмов развития этих реакций (Schaefer H., 1983;
Плеханов Г.Ф., 1990). Разобраться в механизмах не удается из-за противоречий,
возникших между результатами, полученными в реальных условиях (под ЛЭП)
и при экспериментальном моделировании (в установках ПЭП). В
первом случае на животных и человека воздействует ВЭП, а во втором ПЭП.
Налицо неадекватность модели, а когда авторы стремятся понять механизм
патогенного действия ЭП идя от конкретного (факты) к абстрактному (теория),
то неадекватность моделей затрудняет решение этой проблемы.
Таким образом, появление в окружающей среде нового урбоэкологического
фактора привело к возникновению новой экологической проблемы, связанной
с изучением роли этого фактора в экзо- и эндоэкологических взаимосвязях.
Решению этой проблемы и посвящена настоящая диссертация, основные материалы
которой были получены при выполнении автором фрагмента госкомитетовской
темы: "Провести комплекс медико-биологических и природоохранных исследований,
связанных со строительством линий сверхвысокого напряжения 750 - 1150 кВ".
Цель работы - провести, сравнительное изучение структурнофункциональных
изменений в организме и популяции млекопитающих, а также в популяциях,
насекомых и растений при взаимодействии их с техногенными вращающимися
и переменными электрическими полями. Выявленные закономерности использовать
для корректировки современных концептуальных моделей процесса взаимодействия
электрических полей с биосистемами, а также для уточнения эквивалентных
электрических схем биосистем различного уровня организации.
Задачи исследования:
1. Изучить влияние техногенных вращающихся и переменных электрических
полей на структуру и динамику популяции беспородных белых мышей,
насекомых и растений.
2. Изучить влияние техногенных вращающихся и переменных электрических
полей на пре- и постнатальный онтогенез беспородных белых мышей.
3. Выявить реакцию кожи и ее производных, а также внутренних
органов (печени, почек, периферической крови, костного мозга) беспородных
белых мышей длительно находившихся в техногенных вращающихся и переменных
электрических полях.
4. Уточнить концептуальную и электрическую модели взаимодействия
биосистем с электрическим полем.
5. Разработать новые способы и устройства для моделирования процесса
взаимодействия внешних электрических полей с биосистемами различного
уровня организации, а также способы и устройства для измерения электрических
параметров животных и человека.
6. Внести корректировки в современные концептуальные модели процесса
взаимодействия электрических полей с биосистемами и в эквивалентные
электрические схемы биосистем различного уровня организации.
7. Предложить наиболее вероятные механизмы возникновения адаптационных
и патологических реакций в биосистемах различного уровня организации при
их взаимодействии с внешними электрическими полями.
Научная новизна исследования.
Впервые проведено сравнительное изучение влияния техногенных вращающихся
и переменных электрических полей низкой частоты на организм и популяции
млекопитающих, насекомых и растений.
Показано, что вращающееся электрическое поле оказывает более
выраженное неблагоприятное воздействие на структуру популяции животных,
насекомых и растений, а также на структурный, энергетический и информационный
гомеостаз организма животных, чем переменное.
Выявлены в покровных тканях мышей, длительно находившихся во вращающемся
электрическом поле, все основные стадии онкогенеза (усиление пролиферации
росткового слоя эпидермиса, доброкачественные и злокачественные
опухоли производных эпидермиса).
Выявлены признаки ускоренного прохождения постнатального онтогенеза
у мышей, длительно находившихся во вращающемся электрическом поле,
по сравнению с переменным.
Обращено внимание на то, что все клетки, ткани и органы млекопитающих
развиваются и функционируют во вращающемся электрическом поле, генерируемом
сердечной мышцей.
Разработана единая эквивалентная электрическая схема взаимодействия
электрических полей с биосистемами, предложены и обоснованы принципиально
новые механизмы взаимодействия электрических полей с человеком и животными.
Объяснен механизм устойчивости пренатального периода млекопитающих
к воздействию внешних электрических полей и механизм их повреждающего действия
в постнатальном периоде.
Научно-практическое значение работы.
Предложены способы и разработаны устройства для моделирования процесса
взаимодействия электрических полей с организмом животных (патенты N 2005407
и
N 2076621, РФ) и с культурой клеток и тканей (патенты N 2076747 и N
2012326, РФ).
Предложены способ и разработано устройство для определения электропроводности
тела животного и биоткани (а. с. N 1659022 и N 1697737, СССР, положительные
решения на изобретение N 5037521, от 25.09.92).
Разработаны устройства для определения выносливости мышц сгибателей
и разгибателей к статическим нагрузкам (а. с. N 1487865, СССР, рационализаторское
предложение N773 от 17.06.88).
Предложен способ моделирования in vivo основных этапов
развития хронической инфекции (Патент N 2094855,
РФ) и способ моделирования процесса упаковки бактерий в мембранные оболочки
(заявка N 96112447/14 от 18.06.96), для моделирования течения инфекционного
процесса на фоне действия электрического поля.
Положения, выносимые на защиту.
1. Выраженность и направленность реакций популяций млекопитающих, насекомых
и растений на техногенное электрическое поле зависит от его формы и напряженности.
2. Вращающиеся и переменные электрические поля промышленной частоты
напряженностью до 14 кВ/м не оказывают существенного влияния на пренатальный
период онтогенеза мышей, однако они вызывают разнообразные по интенсивности
и часто противоположно направленные адаптационные и патологические реакции
в постнатальном периоде, ускоряют его течение.
3. Наиболее выраженные структурно-функциональные изменения на воздействие
техногенных электрических полей выявлены в покровных тканях и менее выраженные
- во внутренних органах. Реакция органов и тканей на воздействие
электрического поля зависит от его формы и напряженности.
4. Современные концептуальные и электрические модели взаимодействия
биосистем с внешним электрическим полем не учитывают модульный принцип
организации биосистем, и соответствующие этим модулям эквивалентные электрические
схемы, а также форму эндогенного электрического поля организма.
5. Разработанные устройства, для измерения электрических параметров
организма, повысили точность измерений. Проводить измерение удельной электропроводности
организма необходимо сразу после выведения его из электрического поля.
Апробация и внедрение результатов в практику.
Основные положения диссертации доложены на: Всесоюзной научно-практической
конференции в Томском НИИ биологии и биофизики, 1985; на 2-й конференции
изобретателей и рационализаторов "Новое в экспериментальной и клинической
медицине", Новосибирск, 1987; на расширенных совместных заседаниях кафедры
биологии НоТКЗМИ и Отдела экологии Новосибирского НИИ энергетики (1984,
1985, 1986, 1987), а также в отчетах о проведенной НИР для этого института
за 1985 и 1987 годы; на научной сессии посвященная 100-летию
Новосибирска (1994); на научной сессии сотрудников НМИ (Новосибирск,
1995); на пятой и шестой научно-практических конференциях врачей
"Актуальные вопросы современной медицины" (Новосибирск, 1995, 1996); на
2 и 3 съездах физиологов Сибири и Дальнего Востока (Новосибирск, 1995,
1997).
Публикации. По теме диссертации опубликовано 33 работы (включая
3 авторских свидетельства и 5 патентов на изобретения).
Структура и объем диссертации. Диссертация состоит из введения,
4 глав, общего заключения, выводов, списка литературы (содержит 204 отечественных
и 52 иностранных работ). Диссертация изложена на 242 стр. машинописного
текста, содержит 28 таблиц и 38 рисунков (графики, схемы, фотографии).
Автор выражает искреннюю признательность за помощь в работе соавторам
публикаций, а также академику В.П.Казначееву за ценные советы и рекомендации
при подготовке диссертации.
Работа выполнена в Новосибирском медицинском институте (ректор
- доктор медицинских наук, профессор А.В.Ефремов). Отдельные фрагменты
работы выполнены совместно с д.б.н., профессором Р.Ш.Ибрагимовым, к.м.н.
А.П.Петровым, к.м.н. Г.М.Вакулиным.
Материалы и методы исследования
В основу работы положены материалы, полученные автором в 1984 - 1988
г. при выполнении трех научно-исследовательских хоздоговорных работ, ответственным
исполнителем которых он являлся: 1. "Изучение влияния электрических полей
промышленной частоты на функционирование генетического аппарата и резистентность
экспериментальных животных". 2. "Изучение влияния сочетанного действия
электрических полей промышленной частоты на лабораторных животных". 3.
"Изучение изменений цитогенетических и биофизических показателей у лабораторных
животных при действии переменных электрических полей".
Темы выполнялись в составе работ ГКНТ СМ СССР и ВЦСПС по проблеме 074
(охрана труда), задание 02 (электробезопасность), этап 02.06.Д1,Д2 "Провести
комплекс медико-биологических и природоохранных исследований, связанных
со строительством линий сверхвысокого напряжения 750 - 1150 кВ".
Эксперименты проводили в установке, созданной сотрудниками отдела экологии
Сибирского НИИ энергетики (к.т.н. Пучков Г.Г., к.т.н. Каскевич Э.П., инженеры
Волков А.П., Соколов А.Г.). Она моделировала ЭП ЛЭП (Пучков Г.Г., и др.,
1986) и состояла из плоских электродов, высоковольтных трансформаторов
и системы управления и защиты персонала от электрического тока. На одну,
вертикально расположенную пару электродов, подавали высокое напряжение
в противофазе. Другая пара электродов располагалась горизонтально. Нижний
электрод был заземлен, а на верхний подавали переменное напряжение со сдвигом
фазы, относительно первой пары электродов, на 90о. В пространстве
между этими электродами формировалось ВЭП. Над верхним электродом был установлен
дополнительный заземленный плоский электрод. Между ними формировалось ПЭП.
Электропитание установки осуществляли от трехфазной сети переменного тока,
напряжением 380 В.
В качестве основного объекта исследования использовали более 3 тысяч
беспородных белых мышей. Их содержали в диэлектрических клетках (Белкин
А.Д., 1987) по 15 животных на клетку. Клетки размещали в центре рабочих
частей установки, на диэлектрических столах. Эксперименты проводили при
двух уровнях напряженности ЭП - высокой и низкой. Высокая напряженность
для ВЭП составляла - 12,5 кВ/м, вертикальная составляющая и 11,0 кВ/м -
горизонтальная, а для ПЭП - 14,0 кВ/м (вертикальная составляющая). Низкая
напряженность для ВЭП составляла - 6,3 кВ/м, вертикальная составляющая
и 5,6 кВ/м - горизонтальная, а для ПЭП - 7,2 кВ/м (вертикальная составляющая).
Животные находились в ЭП по 22 - 23 часа в сутки в течение 8 - 10 месяцев.
Обследование мышей и забор материала проводили в период с 11 до 13 часов.
Самцов и самок исходных групп, раздельно, помещали в ЭП в возрасте 2,5
- 3 месяцев. Затем их ссаживали вместе на неделю (в расчете 2 самца на
4 самки) для получения потомства. От исходной группы было получено три
последовательных генерации животных первого поколения. Самцов и самок первого
поколения скрещивали для получения второго, а второго - для получения третьего.
Скрещивание контрольных животных осуществляли по этой же схеме. Они находились
в схожих условиях, но вне ЭП. Животных забивали методом церебральной дистракции.
В качестве вспомогательных объектов использовали дикую линию
D.melanogaster. Мух (по 2 самца и 2 самки) содержали в стеклянных пробирках
с питательной средой. Пробирки размещали в рабочих частях установки. Контрольная
группа находилась в схожих условиях, но вне ЭП. Температуру помещения поддерживали
на уровне 23 - 24о С. Получено 4 поколения насекомых (более
6,5 тысяч особей), онтогенез которых протекал в ЭП. Другим объектом был
овес (1800 зерен). Зерна проращивали в чашках Петри, расположенных в рабочих
частях установки.
Забор органов и тканей проводили с учетом рекомендаций Г.Г.Автандилова
(1984). Гистологические препараты готовили по стандартным методикам (Ромейс
Б.,1954; Кисели Д., 1962).
При отработке модели процесса эндоэкологического взаимодействия бактерий
с фагоцитами использовали стандартные электронно-микроскопические методики
и оригинальные модели (Белкин А.Д., 1997, Белкин А.Д., Вакулин Г.М., 1997).
Хромосомные препараты готовили из клеток костного мозга (Дыбан
А.П., Баранов В.С., 1987) и окрашивали в 5% растворе красителя Гимза (рН
- 6,8). Хромосомные нарушения учитывали согласно рекомендациям Л.С.Немцова
(1970), Н.П.Бочкова и др., (1972), Evans (1984). Дифференциальное окрашивание
хромосом проводили по методике (Perry, Tomson, 1984) в модификации И.П.Горлова
(1985).
Окраску ядрышек лимфоцитов периферической крови и подсчет нуклеолярного
коэффициента, проводили по методике Smetana et al., (1976) в модификации
А.Д.Белкина (1990).
При проведении морфометрии использовали рекомендации К.Ташкэ (1980),
Г.Г.Автандилова (1984), а при подсчете мест имплантации и живых плодов
- А.П.Дыбан (1978).
Для выявления реакции генетического аппарата на ЭП применяли
метод анализа фенотипа. Животных обследовали на наличие пороков развития,
проводили массометрию тела и внутренних органов (почка, селезенка, печень).
Двигательную активность мышей изучали при помощи теста открытого поля
(Буркацкая Е.М. и др., 1980), а выносливость мышц конечностей к статическим
нагрузкам - при помощи специальных устройств (Белкин А.Д., 1987, 1989).
Удельную электропроводность тела животных определяли при помощи четырехэлектродного
метода (Меницкий Д.Н., 1959; Ибрагимов Р.Ш., 1984) в специальном устройстве
(Белкин А.Д., 1991), а время наступления трупного окоченения у животных
- при помощи методики Л.И.Танка (1957).
Степень агрессивности животных определяли по наличию на их теле следов
от укусов.
Статистическую обработку вариационных рядов проводили по общепринятым
методикам, при Р < 0,05 (Зайцев Г.А., 1984). Вычисляли среднюю величину
(x), ошибку средней (Sx), среднее квадратическое отклонение (S), коэффициент
вариабельности (Cv). Средние величины сравнива-ли при помощи критерия Стьюдента
(t). При альтернативном распределении статистическую обработку осуществляли
путем сравнения долей по методу Фишера (F).
Результаты морфофункциональных исследований по изучению влияния вращающихся
и переменных электрических полей на беспородных белых мышей, зерна овса
и D.Melanogaster
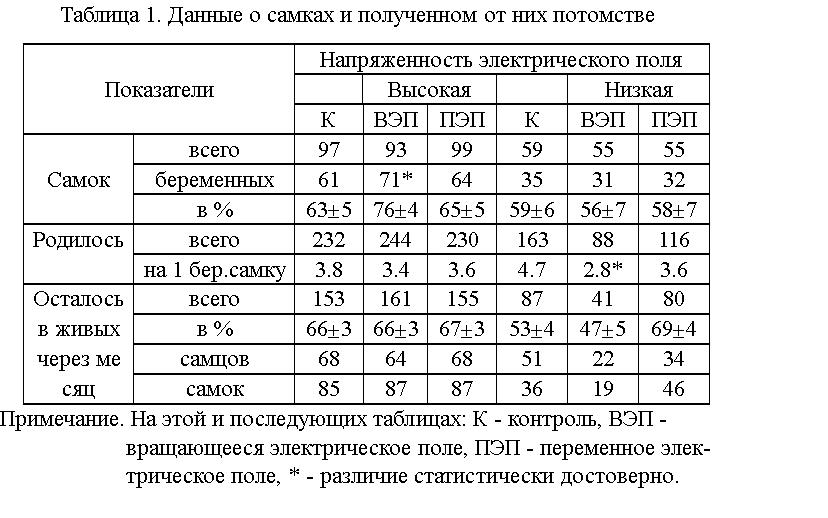
Влияние электрических полей на плодовитость, выживаемость и динамику
веса тела беспородных белых мышей. Изучение влияния ЭП на плодовитость
мышей показало (см. табл.1), что в исходной группе животных, находившихся
во ВЭП высокой напряженности, увеличилась на 13 % доля беременных самок.
Отмечено уменьшение количества новорожденных, приходящихся на одну самку
(на 2 особи), во ВЭП низкой напряженности.
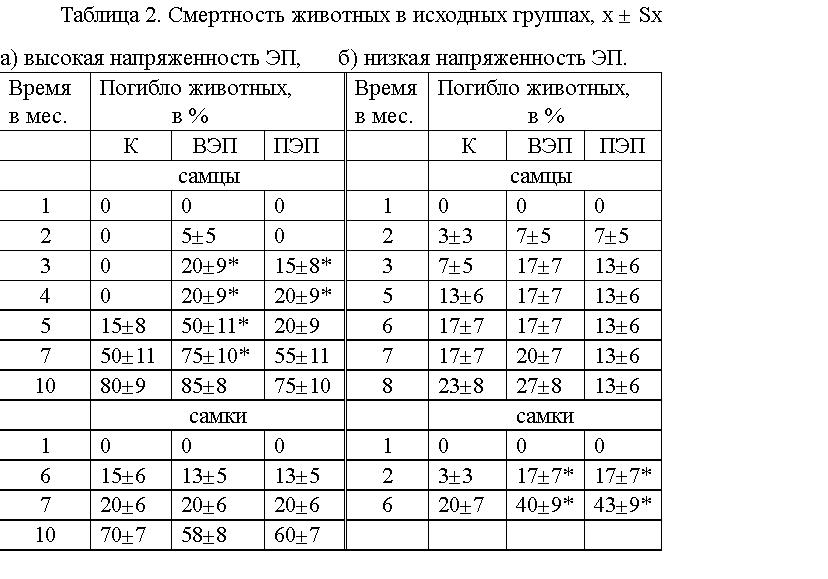
Увеличилась на 5 - 35 % смертность самцов, находившихся в ЭП высокой
напряженности (более выражено во ВЭП), и на 14 - 23 % - самок,
находившихся в ЭП низкой напряженности (см. табл.2), по сравнению
контролем.
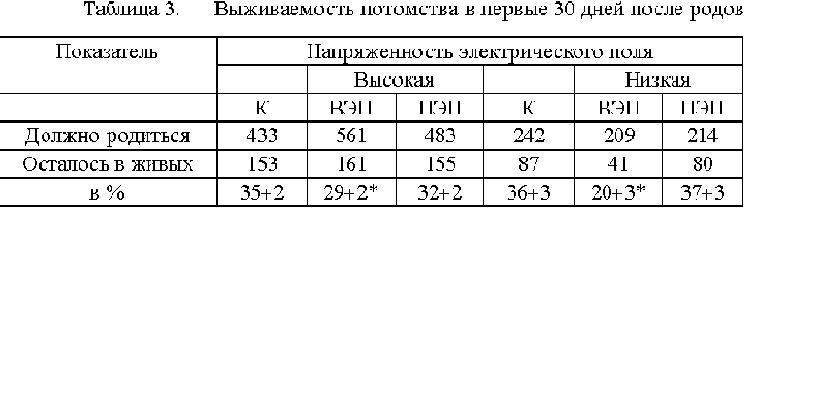
Изучение влияния ВЭП и ПЭП на пренатальный период развития мышей показало,
что ЭП напряженностью до 14 кВ/м не оказывают на него неблагоприятного
влияния. У плодов не выявлено пороков развития, не отмечено и статистически
значимых изменений в до- и постимплантационной смертности, по сравнению
с контролем. Эти данные мы использовали для определения истинной
плодовитости животных. Зная общее количество беременных самок и количество
плодов, приходящихся на одну самку (при их вскрытии),
можно подсчитать каким должен быть приплод, и смертность в раннем постнатальном
периоде (см. табл.3). Как видно из таблицы, в раннем постнатальном периоде,
количество животных, находившихся во ВЭП высокой и низкой напряженности,
уменьшилось на 6 - 16 %.
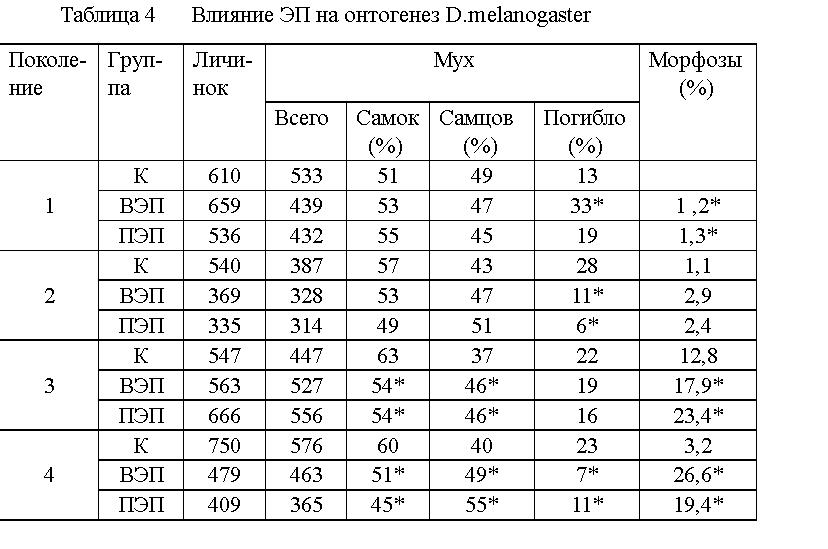
ВЭП оказывает более выраженное влияние на онтогенез и популяционную
структуру D.Melanogaster, чем ПЭП (см. табл.4). У насекомых, находившихся
в ЭП, увеличилось число пороков развития. Наиболее часто встречалась патология
крыльев (их отсутствие или деформация). В 1 поколении смертность насекомых,
находившихся во ВЭП, повысилась на 20 %, а во 2 - 4 - снизилась на 6 -
15 % (во ВЭП и в ПЭП), по сравнению с контролем. Изменилось и соотношение
полов у насекомых, находившихся в ЭП, по сравнению с контролем.
ВЭП замедляет развитие зерновки
овса (в контрольной группе проросло за исследуемый промежуток времени 37
± 2% зерен, во ВЭП - 30 ± 2% и в ПЭП - 36
± 2 %). Различие между контролем и ВЭП статистически достоверно,
а также между ВЭП и ПЭП. Таким образом, на трех группах организмов
показано, что ВЭП оказывает бо-лее выраженное влияние на онтогенез, чем
ПЭП.
Реакция покровных тканей, внутренних органов и системы крови
беспородных белых мышей на воздействие электрических полей. При внешнем
осмотре животных, находившихся в ЭП, и при обзорной микроскопии препаратов,
не выявлено каких-либо патологических изменений в коже и ее производных
(за исключением следов от укусов и опухолей). Это послужило основанием
для проведения морфометрического исследования. Оказалось, что коэффициент
вариации толщины кожи у мышей, находившихся в ЭП, был почти в 2 раза меньше,
чем в контроле (11 и 12% в ВЭП и ПЭП и 21% в контроле). Выявлено также
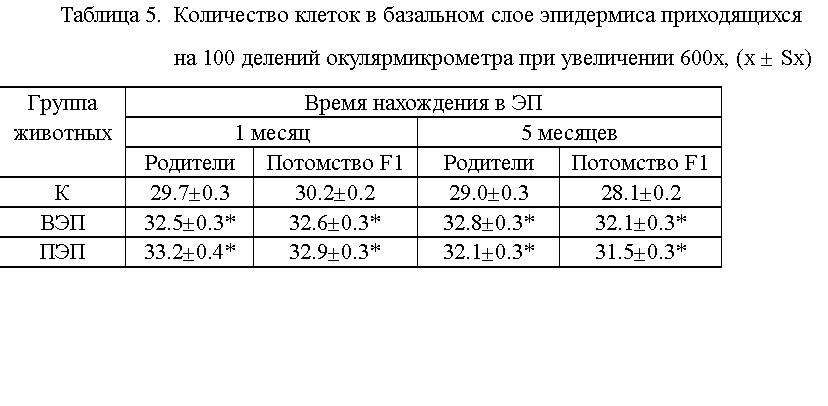
увеличение количества клеток в базальном слое эпидермиса у животных, находившихся
в ЭП, по сравнению с контролем (см. табл. 5). Прибавка составила 2 - 4
тысячи клеток, на 1 мм2.
Среди животных (по 132 особи в каждой группе), находившихся во ВЭП,
выявлена 1 доброкачественная (фиброаденома) и 6 злокачественных (аденокарциномы)
опухолей молочной железы и 1 сосудистая опухоль (гломус-ангиома). В ПЭП
выявлена лишь одна мышь с аденокарциномой молочной железы. В контроле опухолей
не было.
При вскрытии животных и массометрии внутренних органов не зарегистрировано
отклонений в их расположении, окраске, величине, весе. Было решено более
углубленно обследовать только печень и почки. При обзорной
микроскопии препаратов этих органов, также небыло выявлено существенных
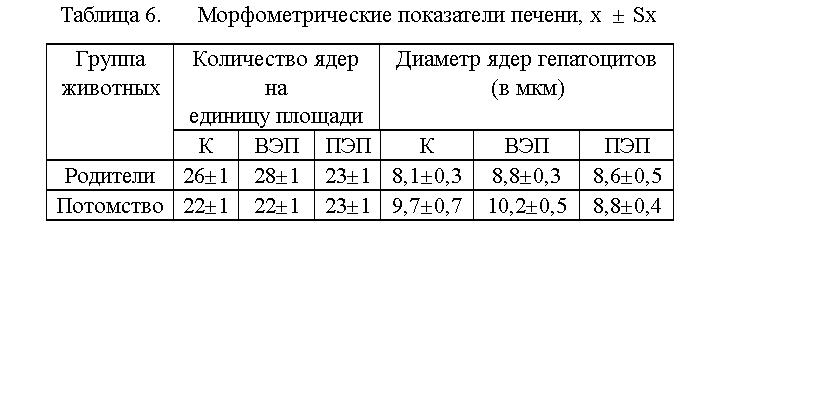
изменений (по сравнению с контролем). Планиметрическое исследование препаратов
печени позволило выявить лишь тенденцию к увеличению (ВЭП) и уменьшению
(ПЭП) числа ядер, приходящихся на единицу площади среза, по сравнению с
контролем (см. табл.6). Различие же между ВЭП и ПЭП, по данному критерию,
было более значимым. Так у животных, находившихся во ВЭП, количество ядер
гепатоцитов, приходящихся на 0,008 мм2, составило 14,1 ± 0,5,
в ПЭП - 13,0 ± 0,5, в контроле - 14,3 ± 0,6, а суммарное число ядер Купферовских,
эндотелиальных и жирнакапливающих клеток, составило для ВЭП
- 14,1 ± 0,9, для ПЭП - 10,4 ± 0,6, в контроле - 12,2 ± 1,0.
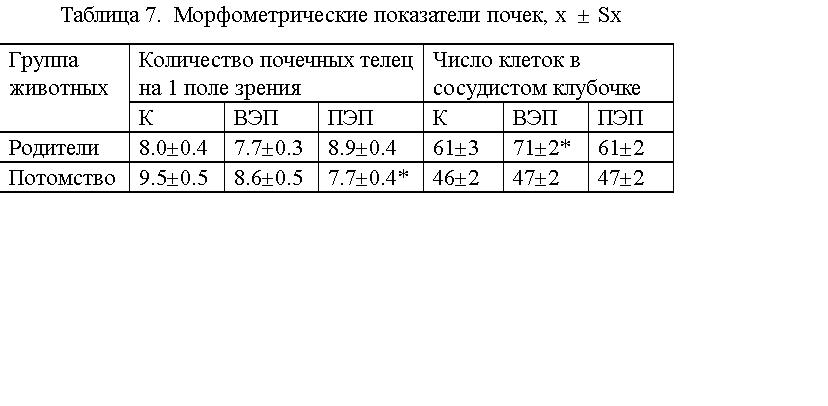
Более выраженные изменения выявлены при морфометрии почек. При планиметрии
(см. табл.7) отмечена тенденция к увеличению количества почечных телец,
приходящихся на единицу площади среза, у животных, находившихся в
ПЭП (исходная группа) и статистически значимое снижение данного показателя
(на 19 %) у потомства, находившегося в ПЭП. Выявлено статистически значимое
увеличение (на 14 %) количества клеток в почечных клубочках у животных,
находившихся во ВЭП (исходная группа). У животных первого поколения сохранялась
лишь тенденция к увеличению данного показателя.
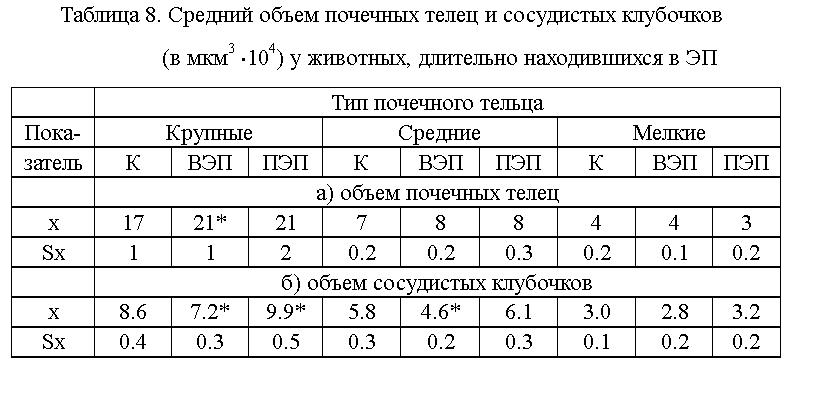
При стереометрии (см. табл. 8) установлено статистически значимое
увеличение (на 20 - 25 %) доли крупных почечных телец у животных, находившихся
во ВЭП. Сосудистые клубочки крупных и средних почечных телец, у животных,
находившихся во ВЭП, были в состоянии спазма, а у животных, находившихся
в ПЭП, - в состоянии дилятации (см. табл.8). Различие между ВЭП и ПЭП статистически
достоверно.
При изучении препаратов крови животных, находившихся во ВЭП, не было
выявлено признаков повреждения клеток, по сравнению с контролем (обзорная
микроскопия и морфометрия). В дальнейшем мы приступили к изучению количественных
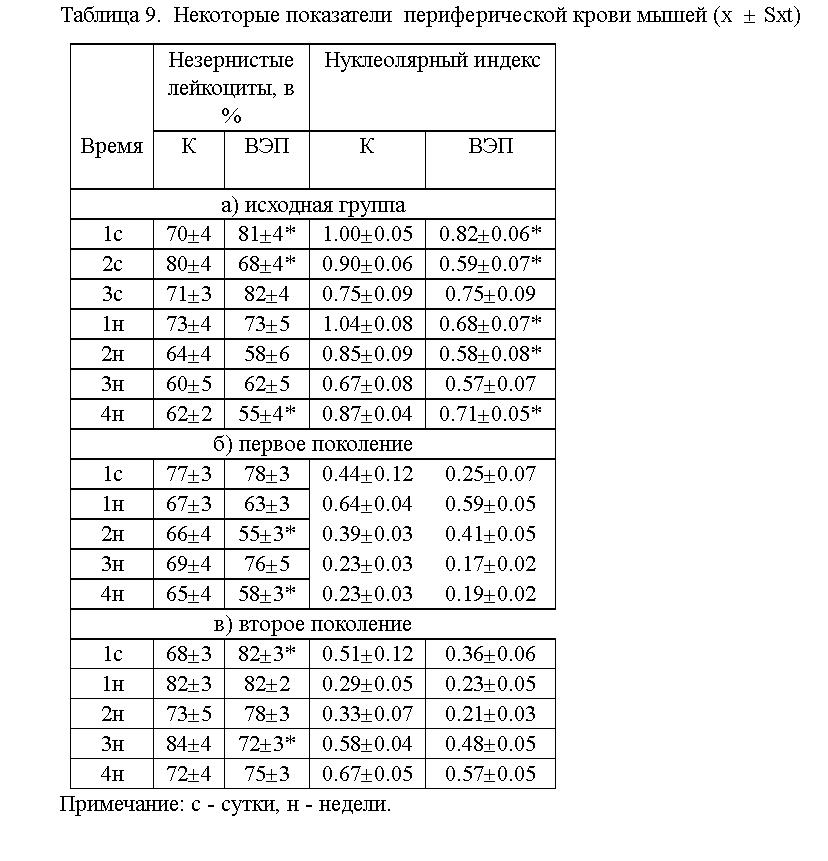
показателей (см. табл.9). Как видно из таблицы клеточный состав периферической
крови слабо прореагировал на воздействие ВЭП.
Учитывая то, что в периферической крови на воздействие ВЭП прореагировали
только лимфоциты, мы попытались оценить их функциональную активность при
помощи нуклеолярного теста (см. табл.9). У животных, помещенных во ВЭП,
уже с первых суток отмечается подавление функциональной активности
лимфоцитов, с одновременным повышением коэффициента вариабельности. У животных
первого и второго поколения, онтогенез которых протекал в ЭП, выявлена
четкая тенденция к снижению зтого показателя (по сравнению с контролем).
Таким образом, в периферической крови, зарегистрированы только функциональные
изменения и то не очень выраженные. Поэтому, при исследовании костного
мозга, было решено ограничиться лишь хромосомным анализом. Обследовали
животных второго поколения, онтогенез которых протекал во ВЭП. В
результате нормальных метафаз в клетках животных, находивших
ся во ВЭП, было 97,6 %, одиночных фрагментов 1 %, разрывов по центромере
0,6 %, полиплоидных клеток 0,8 % (в контроле - 97 %, 1 %, 1,3 % и 0,7 %
соответственно). Не выявив повреждений хромосом, мы приступили к изучению
функциональной активности генетического аппарата клеток костного мозга.
Выявлено статистически значимое увеличение частоты межхроматидных обменов,
у животных первого поколения, постоянно находившихся во ВЭП (15,2% на хромосому),
а также у животных, выведенных из него на сутки (14,8% на хромосому), по
сравнению с контролем (12,8%).
Развитие острой циркуляторной гипоксии на фоне воздействия
вращающегося и переменного электрического поля. Опыты проводили на
модели острой циркуляторной гипоксии печени. Мышь (Balb/C) забивали и вскрывали.
Из печени вырезали бритвой несколько одинаковых кусочков и помещали
их в рабочие части электроустановки (Белкин А.Д., А. с. N 2005407, СССР),
состоящие из 2 чашек Петри (в одной размещены 2 плоских электрода,
а в другой - 4). Пространство между элек-тродами заполняли физраствором.
На два электрода, одной из чашек, подавали переменное напряжение (ПЭП),
а на две пары электродов другой чашки, подавали переменное напряжение со
сдвигом фазы на 90о, относительно друг друга (ВЭП).
Чашки Петри, с находящимися в них кусочками печени, помещали в термостат
(37оС) и воздействовали на них ВЭП и ПЭП в течение 2 часов (плотность
тока 10 мА/см2). Затем кусочки печени фиксировали и готовили
препараты, окрашенные гематоксилином и эозином.
ВЭП вызвало более выраженное повреждение гепатоцитов, чем
ПЭП. Например, ядер с признаками кариорексиса и кариолизиса в контроле
было 23 %, в ПЭП - 30 %, в ВЭП - 34 %. Различие между контролем и ЭП статистически
значимо. Кроме этого, на ЭП прореагировали и эритроциты (пойкилоцитоз),
находящиеся в сосудах печени. В контроле подавляющее число эритроцитов
было круглой формы, редко встречаются деформированные клетки. В ПЭП
большая часть эритроцитов деформирована, а в ВЭП - практически все. Таким
образом, ЭП усугубляют гипоксическое повреждение печени, причем ВЭП оказывает
более выраженное воздействие, чем ПЭП.
Влияние электрических полей на двигательную активность и выносливость
лабораторных животных к статическим нагрузкам. Двигательная активность
(тест открытого поля) мышей, находившихся в ЭП высокой напряженности, была
выше, чем в контроле (45 ± 8 пересечений во ВЭП, 49 ± 7 - в ПЭП и
28 ± 6 - в контроле). Более тщательный анализ полученных результатов показал,
что под воздействием ЭП изменилась структура популяции животных (см. табл.
10).
Выносливость мышц к статическим нагрузкам является одним из основных
показателей работоспособности животных. Мы исследовали выносливость сгибателей
и разгибателей конечностей в специально созданных для этой цели устройствах
(Белкин А.Д., 1989). В результате, у животных, находившихся в ЭП, выявлена
лишь тенденция к снижению данного показателя.
Развитие трупного окоченения у беспородных белых мышей длительно
находившихся в электрических полях. Установлено, что у животных, начиная
с 1,5 месяцев пребывания их во ВЭП, первые признаки трупного окоченения
появлялись через 18,4 ± 1,5 мин., в ПЭП - через 16,4 ± 1,2
мин, а в контроле, через 24,8 ± 1,3 мин.. Подобная картина наблюдалась
и у животных, находившихся в ЭП в течение 3 месяцев (30,3 ± 1,8 мин., 29,1
± 2,5 мин. и 40,2 ± 2,2 мин. соответственно). Различие по сравнению с контролем
значимо и достоверно, при Р < 0,05.
Влияние электрических полей на электропроводность тела беспородных
белых мышей. У мышей, находившихся в ЭП (1 - 5 суток и 8 - 9
месяцев), а также у их потомства, измеряли удельную электропроводность
тела. Она не отличалась от контроля. Однако электропроводность внутренних
органов была снижена у животных, находившихся в ЭП в течение года. Так
удельная электропроводность печени у мышей, находившихся во ВЭП, составила
0,114 ± 0,004 См/м (в контроле - 0,135 ± 0,005), а мышц - 0,124 ±
0,005 и 0,149 ± 0,008 соответственно. Позже автор пришел к выводу,
что ЭП изменяет электропроводность тела только в момент его воздействия.
Было проведено дополнительное исследование. Животных помещали на 2 часа
в ПЭП напряженностью 1 кВ/м. Затем их выводили из установки и, поочередно
(контроль, опыт), измеряли удельную электропроводность их тела на
двух частотах (100 и 1000 Гц). В результате выявлена
более низкая удельная электропроводность тела у животных, находившихся
в ЭП, в ранние сроки после выведения их из ЭП (см. рис.1,2).


Рис. 1. Изменение удельной электропроводности тела мышей
после
выведения их из электрического поля.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ ПРОВЕДЕННОГО ИССЛЕДОВАНИЯ
Особенности реакции биосистем различного уровня организации на воздействие
вращающихся и переменных электрических полей. Изучение влияния
ЭП на плодовитость мышей показало, что в исходной группе животных, находившихся
во ВЭП высокой напряженности, увеличилась (на 13 %) доля беременных самок.
Отмечено уменьшение количества новорожденных, приходящихся на одну
самку (на 2 особи), во ВЭП низкой напряженности. Выявлено увеличение смертности
среди самцов (на 5 - 35 %), находившихся в ЭП высокой напряженности (более
выражено во ВЭП), и среди самок (на 14 - 23 %), находившихся в ЭП низкой
напряженности, по сравнению контролем. Как известно, основными характеристиками
популяции являются ее половой состав и число особей, приходящихся
на единицу территории. Полученные нами данные свидетельствуют о нарушении
структуры экспериментальной популяции животных, находившихся во ВЭП.
Результаты наших опытов показывают, что ЭП напряженностью до 14 кВ/м
не оказывают заметного влияния на пренатальный период. Нами не выявлено
ни одного порока развития у плодов, онтогенез которых протекал в ЭП. До
- и постимплантационная смертность также не отличалась от контроля.
Механизм резистентности пренатального периода онтогенеза млекопитающих,
к воздействию ЭП, невозможно объяснить без эквивалентной электрической
схемы (ЭЭС) беременной самки. Она изображена на рис.2. На ней приняты следующие
обозначения: R1, C1 и R2, C2 - сопротивление и емкость рогового слоя эпидермиса
на входе и выходе наведенного тока, R3, C3 - сопротивление и емкость матки
и околоплодных оболочек, R4 и R5 - сопротивление базального слоя эпидермиса
и кровеносных сосудов, R6 - сопротивление плода, С5 и С6 - емкости между
элементами установки, находящимися под напряжением, и самкой. На схеме
изображены несколько мощных шунтов (R3 - R5), по которым наведенный в беременной
самке, под воздействием ЭП, ток огибает плод, не оказывая
на него воздействия.

Рис. 2. Эквивалентная электрическая схема
беременной самки
Таким образом, нами установлен не только факт резистентности
пренатального онтогенеза к воздействию ЭП, но и объяснен механизм
ее формирования.
Теперь перейдем к обсуждению результатов экспериментов по воздействию
ЭП на постнатальный период онтогенеза. В раннем постнатальном периоде выявлено
статистически значимое снижение (на 6 - 16 %) числа животных, находившихся
во ВЭП, как высокой, так и низкой напряженности. Повышенную гибель новорожденных
можно объяснить каннибализмом (самки поедали новорожденных). Почему ВЭП
провоцирует его, а ПЭП не провоцирует? Ответить на этот вопрос можно только
после анализа и обсуждения данных о поздней постнатальной смертности животных.
Мы следили за динамикой численности исходных групп животных, находившихся
в ЭП, в течение 10 месяцев. Начиная с 3 месяца повысилась на 5 - 35% смертность
самцов (особенно во ВЭП). Мы попытались разобраться в этом феномене. При
обследовании самцов было замечено наличие на их теле следов от укусов (признак
агрессивности). Так за 5 месяцев наблюдения в контрольной группе отмечено
20 ± 4% самцов со следами укусов, во ВЭП - 68 ± 7%, в ПЭП -
43 ± 7%. Различие между контролем и животными, находившимися в ЭП, а также
между животными, находившимися во ВЭП и ПЭП, статистически значимо. Таким
образом, ВЭП оказывает более выраженное влияние на животных, чем ПЭП, активизируя
у них двигательную активность и патологических инстинкты (агрессивность,
каннибализм). Определенную роль в активизации каннибализма могут оказывать
и метаболические сдвиги, возникающие в организме животных под воздействием
ЭП. Нами получены косвенные данные о нарушении энергетического гомеостаза
у животных, находившихся во ВЭП. Начиная с 1,5 - 3 месяцев пребывания мышей
в ЭП, у них, после забоя, трупное окоченение наступало на 9 - 11 минут
раньше, чем в контроле, что указывает на нехватку АТФ в мышцах. К подобным
выводам пришли К.В.Стройкова, Т.И.Беляев (1957).
В опытах на насекомых установлено, что ВЭП оказывает более выраженное
влияние на онтогенез и популяционную структуру D.Melanogaster, чем ПЭП.
У насекомых, находившихся в ЭП, увеличилось число пороков развития
(в 1 поколении на 1,2 - 1,3 %, в 3 - на 5,1 - 10,6 %, в 4 -
на 16,2 - 23,4 %). Наиболее часто встречалась патология крыльев реже -
недоразвитие внутренних органов. В 1 поколении смертность насекомых, находившихся
во ВЭП, повысилась на 20 %, а во 2 - 4 - снизилась на 6 - 15 % (во ВЭП
и в ПЭП) по сравнению с контролем. Изменилось и соотношение полов у насекомых,
находившихся в ЭП, по сравнению с контролем (в 3 - 4 поколений).
Механизм формирования морфозов у насекомых под воздействием ЭП не до
конца ясен. Однако, учитывая наличие у них хитинового покрова, определенную
роль, в развитии патологии, должна играть электрострикция. Т.е. под воздействием
ЭП начинает колебаться хитиновый покров личинки с частотой 50 - 100 Гц.
Эти колебания могут привести к нарушению процессов морфогенеза (Орлов В.М.,
1990).
Нами проведено исследование влияния ЭП на прорастание семян овса. Установлено,
что ВЭП оказывает более выраженное тормозящее воздействие на этот процесс
(всхожесть снизилась на 8 %, по сравнению с ПЭП и контролем).
Таким образом, ВЭП оказывает более выраженное патогенное влияние
на онтогенез и структуру популяций млекопитающих, насекомых
и растений, чем ПЭП.
При гистологическом исследовании не выявлено патологических изменений
в коже (за исключением опухолей) у животных, находившихся в ЭП. В
связи с этим было решено провести морфометрическое исследование. При измерении
толщины эпидермиса кожи у животных, находившихся в ЭП, не было выявлено
статистически значимых изменений данного показателя, по сравнению с контролем.
Однако, коэффициент вариации толщины эпидермиса у животных, находившихся
во ВЭП и ПЭП, был почти в 2 раза меньше, чем в контроле. Как известно,
в популяционной биологии, этот коэффициент используют в качестве одного
из основных критериев для оценки реакции популяции на давление окружающей
среды. Клетки эпидермиса можно рассматривать как своеобразную популяцию.
Поэтому уменьшение коэффициента вариации толщины эпидермиса, свидетельствует
о воздействии на него ЭП. Отмечено также увеличение количества клеток в
базальном слое эпидермиса и роговицы у животных, находившихся во ВЭП и
ПЭП, по сравнению с контролем (на 2 - 4 тысячи на каждый мм2).
У самок, находившихся во ВЭП, выявлены доброкачественные и злокачественные
опухоли молочных желез. В ПЭП выявлена лишь одна самка с опухолью. Полученные
результаты указывают на то, что ВЭП оказывает более выраженное влияние
на кожу и ее производные, чем ПЭП, активизирует спонтанный онкогенез. У
животных выявлены все стадии онкогенеза: предопухоль (повышение количества
клеток в базальном слое эпидермиса), доброкачественная опухоль (фиброаденома
молочной железы) и злокачественная опухоль (аденокарцинома молочной железы).
В отечественной и зарубежной литературе также имеются данные о канцерогенном
влиянии ЭП. Однако, механизм активации канцерогенеза ЭП в этих работах
не рассматривается. Мы считаем что, во время кормления самками мышат, происходит
усиление воздействия на нее ЭП (см рис. 3). На рисунке приняты следующие
обозначения: 1 - 8 упрощенные ЭЭС новорожденных мышат, R1 - R8 - сопротивление
молочных желез, R9 - сопротивление внутренней среды тела самки, R10
и С1 - сопротивление и емкость кожи самки, С2 - емкость между элементом
установки, находящимся под напряжением, и мышами, С3 - емкость между самкой
и элементом электроустановки, находящимся под напряжением.
Для объяснения полученных результатов мы используем три факта: 1. Сопротивление
тела новорожденного мышонка ниже взрослого животного (отсутствие
волосяного покрова и больший процент воды в тканях). 2. Сопротивление соска
мыши в лактационном периоде ниже сопротивления кожи. 3. При захвате
мышонком соска самки образуется низкоомный контакт. Как видно из схемы,
сопротивления и емкости мышат включены в цепь параллельно, а это означает,
согласно закону Ома, что их общее сопротивление снижено.

Рис. 3. Эквивалентная электрическая схема кормящей самки
Таким образом, мышата и кормящая их самка образуют единую электрическую
цепь, подключенную к электроустановке и по ней должен протекать больший
по величине ток, чем через каждое животное в отдельности. В связи с этим,
в области молочных желез должна повыситься плотности тока, по сравнению
с другими участками тела самки. Активации под воздействием ВЭП межхроматидного
обмена в соматических клетках, с одновременным торможением их белоксинтезирующего
аппарата, также могут способствовать онкогенезу.
Известно, что ЭП и ток оказывают влияние на пролиферативные процессы
(Дыхнилкин Ю.В. и др., 1969; Мамонтов С.Г., Иванова Л.Н., 1971; Ткаченко
С.С., Руцкий В.В., 1989). Высказывается мнение, что они могут активизировать
онкогенез (Улащик В.С., 1976; Марков Ю.В., 1992), нарушают ионный гомеостаз
клеток (Улащик В.С., 1976). Таков, по мнению автора диссертации,
механизм стимуляции спонтанного канцерогенеза у самок, находившихся во
ВЭП.
В заключении следует обратить внимание исследователей еще на одну выявленную
закономерность. Она связана с наличием многолетней динамики спонтанного
онкогенеза в популяции животных (Кленицкий Я.С., 1933). Выявленные
нами опухоли распределились по годам следующим образом. В 1985 году,
опухолей у животных не было. В 1986 году выявлена одна опухоль у мыши,
находившейся во ВЭП высокой напряженности. В 1987 году выявлено 6 животных
с опухолями (5 во ВЭП низкой напряженности и одна в СЭП низкой напряженности).
В 1988 году эксперименты не проводились, однако, среди 200
интактных беспородных белых мышей, за этот год, выявлено 10
самок с опухолями молочных желез. Таким образом, внешние ЭП могут оказывать
стимулирующее влияние на спонтанный онкогенез, смещая или растягивая во
времени пик его интенсивности.
При вскрытии животных органы имели обычную величину и окраску. Абсолютный
и относительный вес печени, почек и селезенки у животных, находившихся
в ЭП не отличался от контроля. При обзорной микроскопии препаратов печени
и почек животных, находившихся в ЭП, не выявлено существенных изменений
по сравнению с контролем. Планиметрия выявила лишь тенденцию к увеличению
(ВЭП) и уменьшению (ПЭП) числа ядер, приходящихся на единицу площади среза
печени, по сравнению с контролем. Различие же между ВЭП и ПЭП, по
данному критерию, было более значимым. Основной вклад в эти изменения
внесли Купферовские, жирнакапливающие и эндотелиальные клетки.
Более выраженные изменения выявлены при морфометрии почек. Зарегистрирована
тенденция к увеличению количества почечных телец, приходящихся на
единицу площади среза, у животных, находившихся в ПЭП (исходная группа)
и статистически значимое снижение этого показателя (на 19 %) у их потомства.
Выявлено статистически значимое увеличение (на 14 %) количества клеток
в почечных клубочках у животных, находившихся во ВЭП (исходная группа).
У животных первого поколения сохранялась лишь тенденция к увеличению
данного показателя. Увеличилась на 20 - 25 % доля крупных почечных
телец у животных, находившихся в ЭП (преимущественно во ВЭП).
На следующем этапе исследования мы попытались оценить состояние сосудистых
клубочков. В результате было установлено, что у животных, находившихся
во ВЭП, сосудистые клубочки крупных и средних почечных телец были
в состоянии спазма, а у животных, находившихся в ПЭП - в состоянии дилятации.
Различие между ВЭП и ПЭП статистически достоверно.
Таким образом, под воздействием ЭП изменилась гистоархитектоника печени
и почек. В органах прореагировал на воздействие ЭП их сосудистый компонент.
Отмечен спазм (ВЭП) и дилятация (ПЭП) сосудистых клубочков почек, увеличение
(ВЭП) и уменьшение (ПЭП) числа Купферовских, жирнакапливающих и эндотелиальных
клеток в печени. Ранее, при обсуждении ЭЭС беременной самки, мы отмечали
важную роль шунтов в механизме воздействия ЭП на животных и, в частности,
роль сосудистого шунта в предохранении внутренних органов от воздействия
токов, наведенных в теле животного ЭП. Изменения выявляются в тех
структурах, где согласно ЭЭС должна быть наибольшая плотность наведенного
тока.
Особо следует отметить факт увеличения доли крупных почечных
телец у животных, находившихся в ЭП. Известно (Хэм А., Кормак Д., 1983),
что в почках старых животных, по сравнению с молодыми, преобладают крупные
почечные тельца. Поэтому данный признак мы связываем с влиянием ЭП на процесс
старения (на это указывает и факт появления опухолей у животных,
находившихся во ВЭП). Причем ВЭП оказывает более выраженное влияние на
этот процесс, чем ПЭП. Имеются сообщения о том, что ЭП ускоряют развитие
растительных (Тетюев В.А., Булатов В.А., 1972, 1973; Блонская А.П. и др.,
1985) и животных организмов (Карташов А.Г., Иванова Л.А., 1988;
Карташов А.Г., Мигалкин И.В., 1991; Савельев С.В. и др., 1994). Полученные
факты подтверждают гипотезу о том, что одной из причин акселерации является
электромагнитное загрязнение окружающей среды.
Клеточный состав периферической крови слабо прореагировал на воздействие
ЭП. Подобные изменения у животных, находившихся в ЭП, отмечают и другие
авторы (Schaefer, 1983; Плеханов Г.Ф., 1990). По-видимому, мощные гомеостатические
системы крови и транзитный транспорт через кровь форменных элементов, слабо
реагируют на воздействие ЭП.
Разнонаправленные колебания числа лимфоцитов крови, в первые трое суток
после помещения животных во ВЭП, по сравнению с контролем, свидетельствуют
о том, что ВЭП оказывает модулирующее влияние на течение стресс реакции
(ранее нами отмечено модулирующее влияние ВЭП на спонтанный онкогенез).
Мы считаем, что эта реакция развивается у животных не на воздействие ВЭП,
а на процедуру забора крови. ВЭП, на фоне стресса, оказывало более выраженное
тормозящее влияния на функциональную активность генетического и белоксинтезирующего
аппарата лимфоцитов (на 15 - 37 %). Количество лимфоцитов, в первые трое
суток, колебалось в группе животных, находившихся во ВЭП, противофазно,
по сравнению с контролем.
Таким образом, в периферической крови зарегистрированы только функциональные
изменения, и то не очень выраженные. Поэтому, при исследовании костного
мозга решили ограничиться хромосомным анализом и изучением функциональной
активность генетического аппарата клеток. Обследовали животных второго
поколения, онтогенез которых протекал во ВЭП. Установлено, что частота
хромосомных аберраций в клетках костного мозга животных, находившихся
в ВЭП, не отличалась от контроля.
Не выявив изменений в кариотипе, мы приступили к изучению функциональной
активности генетического аппарата соматических клеток. Обследовали животных
первого поколения, онтогенез которых протекал во ВЭП. Выявлено увеличение
на 2,4 % частоты межхроматидных обменов у животных, находившихся во ВЭП,
по сравнению с контролем. Подобные исследования были проведены Л.А.Поляковой
и А.Г.Карташовым, (1991). Они воздействовали на мышей ПЭП. У молодых животных
частота межхроматидных обменов не отличалась от контроля, у старых - наметилась
лишь тенденция к увеличению этого показателя. По-видимому, ВЭП и ПЭП поразному
воздействуют на функциональную активность генетического аппарата клеток.
Интересно сравнить данные нуклеолярного теста с данными межхроматидного
обмена. Отмеченное снижение функциональной активности белоксинтезирующего
и генетического аппарата лимфоцитов периферической крови у мышей,
находившихся во ВЭП, свидетельствует о блокаде процесса транскрипции и
трансляции. ВЭП как бы настраивает клетку на ускоренное вхождение ее в
митотический цикл (ускоренное старение клетки). Это еще один признак,
свидетельствующий о влиянии ЭП на процесс старения животных.
Двигательная активность (тест открытого поля) у животных, находившихся
в ЭП, была повышена, по сравнению с контролем (на 17 % в ВЭП и 21 % в ПЭП).
Подобный эффект отмечен и Hilmer et al. (1970). Полученные результаты согласуются
с данными о повышении агрессивности у животных, находившихся в ЭП. Все
это указывает на то, что ЭП оказывают воздействие на нервную систему. Имеются
литературные данные, в которых приводятся сведения о нарушении процессов
возбуждения и торможения в коре головного мозга животных, находившихся
в ЭП (Попович В.М., Козярин И.П., 1977; Козярин И.П., Швайко И.И., 1985).
Выносливость животных к физическим нагрузкам является одним из основных
показателей их работоспособности. В результате проведенного обследования
у животных, находившихся в ЭП, выявлена лишь тенденция к снижению данного
показателя, по мере увеличения их возраста, по сравнению с контролем. Существенного
различия между ВЭП и ПЭП не установлено. Таким образом, ЭП оказывают
слабое влияние на этот показатель.
Нами была предпринята попытка косвенно оценить энергетический потенциал
организма мышей, используя методику Л.И.Танка (1957). Она основана на экспериментально
установленном факте, что скорость трупного окоченения зависит от содержания
АТФ в мышцах: чем меньше запас АТФ, тем быстрее оно наступает.
Установлено, что у животных, забитых после 1,5 месяцев пребывания их во
ВЭП, первые признаки трупного окоченения появляются на 6,4 мин. раньше,
чем в контроле, а в ПЭП - на 8,4 мин. Подобный эффект отмечен и у животных,
находившихся в ЭП в течение 3 месяцев (9,9 и 11,1 мин. соответственно).
Различие по сравнению с контролем значимо и достоверно при Р < 0,05.
Обсуждая ЭЭС биоткани, мы отмечали, что при протекании наведенных токов
через биоткань должен нарушаться ионный гомеостаз клеток. Его восстановление,
как известно, осуществляется за счет расхода АТФ. В.С.Улащик (1976) показал,
что при протекании через организм гальванического тока плотностью 0,3 мА/см2
в тканях снижается концентрация АТФ, а Н.М.Громыко и др., (1991) приводят
данные о том, что у животных, находившихся в ЭП, выявляется тканевая гипоксия.
Таким образом, нами получен косвенный признак, указывающий на снижение
АТФ в тканях животных находившихся в ЭП. Кроме этого, при проведении эксперимента
по воздействию ЭП на течение острой циркуляторной гипоксии печени, выявлено,
что ЭП усугубляют ее течение (особенно ВЭП). Снижение выносливости
животных к статическим нагруз-кам также указывает на неблагополучие в энергетическом
гомеостазе.
Учитывая то, что при взаимодействии животного с ЭП его тело становится
элементом электрической цепи, мы решили проследить за изменением удельной
электропроводности тела животного в процессе эксперимента. Оказалось, что
нарушенная под воздействием ЭП электропроводность тела быстро восстанавливается
после выведения животных из ЭП. Поэтому при проведении подобных измерений
необходимо учитывать этот эффект.
Обсуждение концептуальной и электрической моделей взаимодействия
человека с электрическим полем. Рассмотрение процесса взаимодействия
человека с ЭП мы начнем с составления эквивалентной электрической схемы
(ЭЭС) этого взаимодействия, так как без адекватной ЭЭС невозможно разобраться
в механизме действия ЭП. Составляя ЭЭС взаимодействия человека с токонесущим
элементом электроустановки, в методических и практических целях,
следует разбить ее на три схемы: мега-, макро- и микро-ЭЭС. Каждая из схем
предназначена для решения определенного круга задач. Первая - преимущественно
для электротехнических задач, вторая - для задач, связанных с электробезопасностью,
третья - для задач, связанных с вредностью изучаемого фактора (см. Рис.
4).
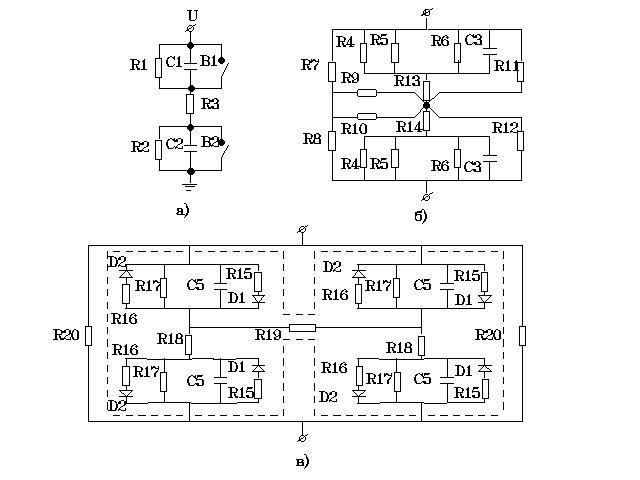
Рис. 4. Эквивалентные электрические схемы взаимодействия человека
с эле-
ментом электроустановки, находящимся под напряжением
(а - мега-ЭЭС, б - макро-ЭЭС, в - микро-ЭЭС)
Вначале рассмотрим мега-ЭЭС. Основные варианты взаимодействия человека
с электроустановкой хорошо известны (Долин П.А., 1984; Манойлов В.Е., 1985;
Удод Е.И., 1986; Барг И.Г., Полевой С.В., 1989). Исходя из этих работ,
можно составить мега-ЭЭС взаимодействия фазного напряжения элемента электроустановки
с человеком (см. рис. 4 а). На этой схеме приняты следующие условные обозначения:
U - фазное напряжение, С1 и С2 - емкость человека относительно фазы и земли,
R1 и R2 - сопротивление воздуха между обкладками конденсатора С1 и С2,
R3 - сопротивление тела человека, В1, В2 - выключатели. Мега-ЭЭС моделирует
четыре основных типа взаимодействия человека с фазным напряжением:
1. Если выключатели В1 и В2 замкнуты, то моделируется короткое
замыкание. Человек попадает под полное фазное напряжение.
2. При разомкнутом выключателе В1 и замкнутом В2 моделируется взаимодействие
заземленного человека с фазным напряжением.
3. При замкнутом выклю-чателе В1 и разомкнутом В2 моделируется выход
человека на фазу.
4. При разомкну-тых выключателях человек полностью изолирован
от токонесущих частей электроустановки и земли.
Данная мега-ЭЭС для инженеров-электриков ясна. Имеются предварительные
расчеты ее отдельных звеньев (Долин П.А., 1984; Манойлов В.Е., 1985; Удод
Е.И., 1986; Барг И.Г., Полевой С.В., 1989, Schaefer H., 1983). Чтобы окончательно
разобраться в данной проблеме, необходимо расшифровать более тонкие механизмы
взаимодействия ЭП с биосистемами и сделать более точные расчеты. Анализ
мега-ЭЭС взаимодействия человека с фазным напряжением позволяет наметить
несколько вариантов механизма этого взаимодействия. Первый вариант связан
с наведением на поверхности тела переменного потенциала (кулоновское взаимодействие),
второй - с деформацией диэлектрика (кожи) под воздействием ЭП (электрострикция),
третий - с наведенными в биообъекте токами.
Из вышеизложенного вытекает очевидный факт, что без адекватной ЭЭС
тела человека разобраться в этих механизмах невозможно. При создании макро-
и микро-ЭЭС необходимо учесть анатомо-гистологические и цитологические
особенности строения человека и животных. Прежде всего, на макроскопическом
уровне, имеется мощный шунт (пищеварительный тракт) и несколько участков
поверхности тела с высокой электропроводностью (глаза, носовая полость,
мочеполовые отверстия). Все эти образования являются входными воротами
для наведенных токов. Их влияние необходимо учитывать при изучении процесса
взаимодействия ЭП с человеком и при создании ЭЭС. Например, сопротивление
постоянному току, между передними и задними конечностями мыши, составляет
1 мОм, а между начальным и конечным отделом пищеварительного тракта - 10
кОм (в 100 раз меньше). Сопротивление же переменному току (50 Гц, 5,8 В)
составило соответственно - 72,5 и 2,4 кОм (в 30 раз меньше). На рис. 4
б изображена макро-ЭЭС тела человека. На ней приняты следующие условные
обозначения: С3 и R4 - емкость и сопротивление рогового слоя эпидермиса,
R5 и R6 - сопротивление потовых желез и биоактивных точек, R7 и R8 - сопротивление
начального и конечного отделов пищеварительного тракта, R9 и R10 - сопротивление
желчных протоков и протоков поджелудочной железы, R11 и R12 - сопротивление
глаз и протоков мочеполовой системы, R13 и R14 - сопротивление внутренних
органов и тканей. Все сопротивления комплексные.
Теперь нам осталось рассмотреть микро-ЭЭС (см. рис. 4 в). При ее создании
учтено слоистое строение тканей, ионная асимметрия клеточных мембран и
избирательная проницаемость их для различных ионов. На ней приняты следующие
условные обозначения: D1 и D2 - натриевые и калиевые каналы клетки, обладающие
односторонней проводимостью, R15 и R16 - сопротивления натриевых и калиевых
каналов, R17 и С5 сопротивление и емкость мембраны клетки, R18 - сопротивление
цитоплазмы, R19 - сопротивление межклеточного щелевого контакта, R20 -
сопротивление межклеточной среды. Пунктиром обозначена граница клетки.
Все схемы совместимы и составляют единую ЭЭС. Она послужила основой
для создания концептуальной модели взаимодействия человека с ЭП. Энергия,
заключенная в ЭП, является главным двигателем всех процессов, протекающих
в электрической цепи, сформированной между элементом электроустановки,
находящимся под напряжением, и поверхностью земли. Мега-ЭЭС можно представить
в виде огромного конденсатора. Его обкладками являются токонесущая часть
электроустановки и поверхность земли. Атмосферный воздух, заполняющий его
пространство, играет роль диэлектрика. Так как он не идеальный диэлектрик,
то через этот конденсатор протекает не только ток смещения, но и ток проводимости.
Человек, попадая в межэлектродное пространство конденсатора, изменяет его
свойства, так как с одной стороны он является диэлектриком (роговой слой
эпидермиса кожи), а с другой - вносит в это пространство значительный объем
вещества с высокой электропроводностью (органы и ткани тела). ЭП поляризует
эпидермиса и наводит в организме ток проводимости и смещения. Поляризация
эпидермиса сопровождается его механическими колебаниями (электрострикция).
Между током смещения и проводимости имеется сдвиг фаз.
Итак, в пространстве между элементом электроустановки и человеком,
действуют два фактора: ЭП и электрический ток. От их параметров (напряженности
ЭП, частоты и силы тока) зависит состояние человека и окружающей
среды.
Четыре основных механизма взаимодействия человека с фазным напряжением
можно объединить в два: опасный для жизни и неопасный. Опасными для жизни
являются: механизм, сопровождающийся пробоем воздушного промежутка между
человеком и элементом электроустановки, находящимся под напряжением
и касание заземленного человека токонесущих частей. Эти варианты хорошо
изучены (Долин П.А., 1984; Манойлов В.Е., 1985). Установлены опасные (100
мА), безопасные (0,1 мА) и пороговые (1 - 5 мА) значения тока. Несмотря
на то, что этим вариантом механиз-ма действия уже много лет занимается
целая отрасль - электробезопасность, выявле-ны лишь наиболее грубые изменения
в организме животных и человека, пораженных электрическим током. Сам же
механизм до конца не раскрыт. Что же происходит в организме человека при
протекании тока менее 0,1 мА, большинство ученых, работавших в области
электробезопасности, не интересовало. Нет поражения человека, нет и проблемы.
Кроме этого морфологи не выявляли никаких специфических изменений в органах
и тканях у лиц, погибших от электрического тока (Орлов А.Н., Саркисов М.А.,1977).
Опасный для жизни вариант механизма действия ЭП отличается от безопасного
тем, что в первом случае поражающий фактор (ток) воздействует на ограниченный
участок тела человека, а во втором - на весь организм, так как он является
частью среды, по которой протекают токи и силовые линии ЭП. Объединяет
эти два варианта электрический ток. В первом случае он достигает больших
величин (опасных), во втором - небольших (безопасных). При этом возникает
вопрос: Что же происходит в организме человека при протекании через него
токов ниже опасных величин? Основываясь на макро-ЭЭС можно описать макромеханизм.
Человек, становясь элементом мега-ЭЭС, попадает под воздействие всех
процессов, протекающих в ней. Так как тело человека состоит из неэлектропроводной
оболочки и содержимого, заключенного в ней с высокой электропроводностью,
то, вытеснив своим объемом часть объема воздуха, тело человека, обладающее
большей электропроводностью, чем воздух, начинает притягивать к себе силовые
линии ЭП и токи. Этому способствуют и области с высокой электропроводностью
на поверхности тела (глаза, ротовая и носовая полости, анальное и
мочеполовые отверстия). Часть из них шунтирует токи, протекающие через
организм (пищеварительный тракт), остальные играют роль входных ворот.
Через эти области протекают токи проводимости и смещения. Что же касается
электро-стрикции, то она формирует один из дополнительных механизмов, который
может способствовать изменению проницаемости эпидермиса. Кроме этого, наведенные
на коже, под воздействием ЭП, потенциалы изменяют картину естественных
потенциалов эпидермиса и могут нарушать информационные потоки между участками
кожи и внутренними органами.
Микромеханизм формируется за счет особенностей микро-ЭЭС и путей токов,
протекающих через покровные ткани и внутренние органы. Как отмечалось
выше, эпидермис обладает рядом особенностей, способствующих протеканию
через него тока. Входными воротами на микро уровне являются клетки биоактивных
точек и потовых желез. Сопротивление этих участков кожи намного
ниже, чем остальных. Так как в ЭЭС они расположены параллельно, то их удельные
электропроводности суммируются, и общая электропроводность эпидермиса будет
выше электропроводности отдельной биоактивной точки или потовой
железы. Далее ток протекает через ионные каналы мембран клеток эпидермиса.
Сосредоточив основное внимание на носителях тока - ионах, и учитывая асимметрию
их распределения по обеим сторонам клеточной мембраны и селективную избирательность
ионных каналов, можно представить следующий механизм протекания тока через
клетку. Известно, что снаружи клетки преобладают ионы натрия, а внутри
- калия (для простоты рассуждений мы не рассматриваем другие ионы).
Электрохимический градиент натрия направлен внутрь клетки, а калия наружу.
В связи с этим, входящий ток будет связан с ионами натрия, а выходящий
- с ионами калия. Поэтому ток будет втекать в клетку через натриевые каналы
(D1) и ионы натрия начнут поступать в клетку, а вытекать через калиевые
каналы (D2) выводя ионы калия в межклеточное пространство. В результате
произойдет нарушение ионного гомеостаза клетки. Для его восстановления
она должна затратить АТФ (удалить лишние ионы натрия и ввести внутрь недостающие
ионы калия). Этому механизму способствует и тот факт, что ток частотой
50 Гц вызывает наибольшие колебания ионов по сравнению с другими частотами
(Долин П.А.1984). Следует иметь в виду, что сопротивления ионных каналов
(как и биоактивных точек) включены в ЭЭС параллельно и поэтому общая
электропроводность клетки выше электропроводности одиночного ионного канала.
Следующее звено механизма связано с распределением ветвей тока между
клетками и межклеточной средой. Чем более выражена межклеточная среда у
данного типа ткани, тем более выражен ее шунтирующий эффект. Одной из особенностей
эпидермиса кожи, по сравнению с другими тканями, является слабая выраженность
межклеточной среды. Клетки эпидермиса достаточно прочно скреплены межклеточными
контактами. Но наряду с плотными контактами эти клетки обладают и щелевидными.
Через них идет свободный обмен ионами между клетками ткани. Поэтому ток,
проникший внутрь клеток, не будет из них выходить, а начнет шунтироваться
через межклеточные щелевидные контакты (R19) по слою эпидермиса. Таким
образом, имеется еще один шунт, защищающий внутреннюю среду организма от
наведенных токов. Если ток преодолевает барьер эпидермиса, то он замыкается
в новом шунте - кровеносных сосудах. Преодолев этот шунт, ток оказывает
влияние на клетки внутренних органов и тканей. Учитывая то, что все процессы
жизнедеятельности клеток и тканей, а также органов и систем связаны с ЭП
и токами, создаваемыми этими образованьями, протекание через них дополнительных
токов должно сказываться на информационном гомеостазе (тканевом и клеточном).
Нами выявлены признаки нарушения этих видов гомеостаза (увеличение количества
клеток в базальном слое эпидермиса кожи и роговицы, опухоли молочных желез,
увеличение частоты сестринских межхроматидных обменов в клетках у мышей,
длительно находившихся в ЭП). Таким образом, ЭЭС взаимодействия человека
с элементом электроустановки, находящимся под напряжением, и механизмы,
определяемые этими схемами и особенностями строения тела человека, позволяют
дать наиболее полное словесное описание данного процесса и создать его
математическую модель. Наиболее перспективным при разработке этой модели
является путь от описания и расчета наиболее общих процессов к частным,
от мега- к микро уровню. Самой первой задачей является установление границ
колебания показателей (плотностей токов, напряженности ЭП), исходя
из реальной ситуации, на уровне мега-, макро- и микро-ЭЭС. Затем необходимо
уточнить колебания отдельных показателей на уровне каждой из схем и выявить
первичные эффекты, напрямую связанные с воздействующими факторами. На следующем
этапе необходимо проследить за вторичными, третичными и т.д. эффектами,
выявив взаимосвязи между ними. Подобная работа для мега-ЭЭС уже частично
проведена (Долин П.А., 1984; Удод Е.И., 1986; Барг И.Г., Полевой С.В.,
1989), что касается макро- и мик-ро-ЭЭС, то эту работу предстоит еще сделать.
Экологическая значимость природных и антропогенных ЭП. Самыми
загадочными и не до конца исследованными являются механизмы слабых
экологических взаимодействий. Огромную роль в этих процессах играют ЭМП.
Если для ихтиофауны, в настоящее время, никто не будет отрицать экологическую
значимость ЭМП (Протасов В.Р., и др., 1982; Басов Б.М., 1985; Шарова Л.В.,1987;
Савельев С.В., и др., 1994), то для наземной фауны и флоры их роль не так
очевидна. Успехи в изучении роли ЭМП в экологических взаимосвязях с ихтиофауной
позволили В.Р.Протасову и др., (1982) выделить новое направление в экологии
- электроэкологию. Ее основной задачей является изучение электрических
взаимоотношений в живой природе. Учитывая, что экологию часто подразделяют
на экзоэкологию (экологию внешней среды) и эндоэкологию (экологию внутренней
среды организма), можно выделить экзоэлектроэкологию и эндоэлектроэкологию.
Сложнее обстоит дело с изучение роли ЭМП в экологических взаимосвязях
наземных животных. Делались неоднократные попытки выяснить их экологическую
значимость (Пресман А.С., 1974; Дубров А.П., 1974).
Развитие системы энергетики и ввод в эксплуатацию ЛЭП высокого и сверхвысокого
напряжения поставили перед исследователями вопрос об экологической значимости
ЭП промышленной частоты. Во всех экономически развитых странах началось
интенсивное изучение влияния электрических, электромагнитных и магнитных
полей на организм человека и животных. При проведении этих исследований,
как сообщалось выше, ученые не обратили внимания на форму ЭП генерируемого
ЛЭП, считая его переменным, в то время как оно было вращающимся.
Таким образом, незаметно для исследователей, всего за каких-то 50 лет сменился
электромагнитный фон окружающей среды. В экзоэлектроэкологии произошел
переход от статических и переменных к вращающимся ЭП. Событие совершилось,
и никто не знает его последствий. Установление этого факта помогло автору
диссертации обратить внимание на форму ЭП определяющего эндоэлектроэкологические
взаимодействия. Положение в эндоэлектроэкологии оказалось схожим с экзоэлектроэкологией.
Несмотря на то, что было известно, что сердце генерирует вращающееся ЭП
(на этом основан метод векторкардиографии) никто из исследователей,
занимающихся проблемами физиологии и электромагнитной биологии, не обратил
на это внимание. В результате, был упущен важный факт - все клетки и органы
человека и животных функционируют во вращающемся ЭП. Установление этого
факта позволяет сделать предположение о том, что биополем, которое искал
А.Г.Гурвич, является эндогенное ВЭП. Таким образом, можно считать, что
в эндоэлектроэкологических взаимосвязях важную роль должны играть ВЭП.
В экзоэлектроэкологии, до последнего времени, преобладали СЭП и ПЭП. Такое
различие между эндо- и экзоэлектроэкологией сложилось в результате длительной
эволюции и, по-видимому, было необходимо для более четкого отделения
внутренней полевой среды организма от внешней. И вот, за короткий промежуток
времени, исчезла эта полевая отграниченность. В связи с тем, что окружающая
среда заполнена электросмогом и радикально, в ближайшем будущем, изменить
обстановку невозможно, особую роль приобретают теоретические исследования
и целенаправленные эксперименты по выявлению и изучению отдельных звеньев
слабых экологических взаимодействий. Поэтому одной из задач работы была
разработка способов и устройств для моделирования процесса взаимодействия
ЭП с биосистемами (патенты РФ N 2005407, N 2012326, N 2076621) и
для определения их состояния (а. с. РФ N 1487865, N 1659022, N 1697737,
патенты РФ N 2006207, N 2043792).
ОБЩЕЕ ЗАКЛЮЧЕНИЕ
Таким образом, результаты сравнительного изучения влияния вращающихся
и переменных ЭП на лабораторных животных, насекомых и растения позволяют
сделать вывод о том, что эти два вида ЭП различаются не только по своей
форме, но и по механизму действия. Нами получены данные о разнонаправленных
адаптивных и патологических реакциях популяции животных, насекомых, растений,
а также органов, тканей и клеток животных, на воздействие этих двух видов
ЭП. Эти данные послужили основой для проведения теоретических исследований,
направленных на преодоление противоречий, возникших при изучении "слабых"
эндо- и экзоэкологических взаимодействий, вызванных невнимательным отношением
исследователей к методологическим и методическим аспектам данной проблемы.
Например, недоучет формы ЭП приводил к смешению результатов экспериментов
по воздействию на животных вращающихся и переменных ЭП, что затрудняло
анализ и интерпретацию полученных данных и, в конечном счете, создавало
непреодолимые трудности при расшифровке механизма действия ЭП. Подавляющее
большинство авторов, проводивших исследования по изучению влияния ЭП на
биосистемы, не имели четкого представления об электрической схеме этого
взаимодействия, что не способствовало раскрытию его механизма. Наше исследование
восполнило многие пробелы и заложило основы для окончательной расшифровки
механизмов взаимодействия ЭП с биосистемами.
В Ы В О Д Ы
1. Вращающееся электрическое поле - новый урбоэкологический фактор,
который формируется в городской среде за счет первичных (линии
электропередачи, установки трехфазного тока и электромагнитного излучения)
и вторичных (отраженные электромагнитные поля) источников. Его появление
приводит к нарушению эволюционно сложившегося полевого отграничения внутренней
среды организма человека и животных (вращающееся электрическое поле) от
внешней (статические и переменные электрические поля).
2. Показано, что вращающееся электрическое поле вызывает более выраженные,
чем переменное, изменения в популяции млекопитающих, характеризующиеся
повышением их агрессивности (число животных с признаками укусов на теле
увеличилась во вращающемся электрическом поле на 48,3 %, а
в переменном - на 22,9 %, по сравнением с контролем) и смертности (на 8
- 16 %). У насекомых вращающееся электрическое поле увеличивает на 20%
смертность в 1 поколении и снижает на 6 - 15% во 2 - 4 поколениях. Частота
пороков развития у насекомых, находившихся в электрических полях,
нарастает от 1, к 4 поколению превышая контрольные цифры. У растений снижает
всхожесть овса на 8 %.
3. Вращающееся и переменное электрические поля вызывают нарушения тканевого
и органного гомеостаза у млекопитающих: Увеличилось (на 2 - 4 тысячи на
1 мм2) количество клеток в ростковом слое эпидермиса и роговицы
мышей, подвергнутых воздействию электрических полей. У самок, находившихся
во вращающемся электрическом поле, развились злокачественные (аденокарциномы)
и доброкачественные (фиброаденомы) опухоли молочных желез. В почках животных,
находившихся во вращающемся электрическом поле, возросла на 20 - 25% доля
крупных почечных телец. Выявлена разнонаправленная реакция сосудистых клубочков
нефронов на воздействие электрических полей - спазм во вращающемся электрическом
поле и делятация - в переменном. В клетках периферической крови
животных находившихся во вращающемся электрическом поле понижается на
15 - 37% активность белоксинтезирующего аппарата (по данным нуклеолярного
индекса), а в клетках костного мозга - активизируется на 2,4 % интенсивность
межхроматидных обменов.
4. Жизнь в техногенных электрических полях приводит к сокращению ее
продолжительности, за счет повышения агрессивности самцов друг против друга
(животных со следами укусов на теле было на 48 % больше во вращающемся
электрическом поле и на 23 % - в переменном, по сравнению с контролем)
и самок против своего потомства (за счет каннибализма погибло во вращающемся
электрическом поле на 6 - 16 % больше новорожденных, чем в переменном и
контроле).
5. В организме животных, длительно находившихся во вращающемся электрическом
поле, выявлены признаки преждевременного старения клеток (увеличение количества
клеток в ростковом слое эпидермиса, уменьшение активных ядрышек в лимфоцитах),
органов (увеличение в почках доли крупных почечных телец) и организма (активизация
спонтанного онкогенеза).
6. Предложенные концептуальная модель и единая эквивалентная электрическая
схема обосновывают три основных механизма взаимодействия внешних электрических
полей с биосистемами: а) мегамеханизм (за счет тока проводимости и кулоновского
взаимодействия); б) макромеханизм. Формируется за счет путей прохождения
тока проводимости и смещения через участки поверхности тела с высокой электропроводностью
(глаза, носовая и ротовая полости, анальное отверстие, потовые железы
и биоактивные точки). Желудочно-кишечный тракт шунтирует токи (его сопротивление
переменному току в 30 раз меньше, чем сопротивление кожи), а биоактивные
точки и потовые железы резко снижают сопротивление кожи протекающему току,
так как их сопротивления включены в электрическую цепь параллельно; в)
микромеханизм. Он формируется за счет путей прохождения электрического
тока через клетку. При этом необходимо учитывать ионную асимметрию наружной
клеточной мембраны (электрохимический градиент) и то, что вход тока
в клетку и его выход должен осуществляться за счет разных ионов.
7. Механизм невосприимчивости пренатального периода млекопитающих к
электрическим полям должен обеспечиваться за счет биологических экранов
материнского организма (покровные ткани, матка) и плода (околоплодные оболочки).
Наличие этих экранов указывает на важную роль эндогенного вращающегося
электрического поля в гисто- и органогенезе.
8. При кормлении самкой новорожденных мышат происходит объединение
их тел в единую систему, на которую и воздействует внешнее электрическое
поле. Поэтому эквивалентная электрическая схема каждого новорожденного,
при кормлении, подключена к телу самки, через соответствующую молочную
железу. Это соединение должно способствовать развитию опухолей молочных
желез у кормящих самок, так как ток, наведенный в телах новорожденных электрическим
полем, протекает через молочные железы самки.
СПИСОК РАБОТ, ОПУБЛИКОВАННЫХ ПО ТЕМЕ ДИССЕРТАЦИИ
1. Белкин А.Д. Влияние вращающегося электрического поля высокой напряженности
на лабораторных животных// Тр. ин-та/ Новосиб. гос. мед. ин-т. - 1985.
- Т. 122. - С. 75-77.
2. Белкин А.Д. Анализ хромосомных нарушений в соматических клетках
у мышей, длительно находившихся в электрическом поле промышленной частоты//
Тр. ин-та./ Новосиб. ордена Трудового Красного Знамени мед. ин-т. - 1987.
- Т. 127. - С. 68-69.
3. Белкин А.Д. Клетка для содержания лабораторных животных с устройством
для регистрации двигательной активности// Новое в экспериментальной и клинической
медицине: Тез. докл. ко 2-й конф. Изобретателей и рационализаторов. 11
декабря 1987 г. - Новосибирск, 1987. - С.
108-109.
4. Белкин А.Д. Устройство для определения выносливости лабораторных
животных к статическим нагрузкам и выработки условных рефлексов//Новое
в экспериментальной и клинической медицине: Тез. докл. ко 2-й конф. изобретателей
и рационализаторов. 11 декабря 1978 г. - Новосибирск, 1987. - С. 112-113.
5. Белкин А.Д. Об одном из методологических подходов в экспериментальной
биологии и медицине// Тр. ин-та/ Новосиб. ордена Трудового Красного
Знамени мед. ин-т. - 1989. - Т. 133. - С. 88-92.
6. Белкин А.Д., Ибрагимов Р.Ш. Влияние внешних электрических полей
на электропроводность внутренних органов// Бюл. Сиб. отд-ния
АМН СССР. - 1989. - N 2. - С. 107-109.
7. Белкин А.Д., Матросова В.Ю., Крылова М.И. Влияние вращающихся
электрических полей промышленной частоты на лабораторных животных//Бюлл.Сиб.
отд-ния АМН СССР.- 1990.- N2. - С.80-83.
8. Белкин А.Д., Петров С.А. Влияние электрических полей промышленной
частоты на спонтанное возникновение опухолей у лабораторных животных//
Бюлл. Сиб. отд-ния АМН СССР.- 1990. - N3,4. - С. 89-91.
9. Белкин А.Д. Устройство для измерения электропроводности лабораторных
животных. А.с. N 1659022. Опубл. в Б.И. N 24, 1991.
10. Белкин А.Д. Устройство для измерения электропроводности биоткани.
А.с. N 1697737. Опубл. в Б.И. N 46, 1991.
11. Белкин А.Д. Реакция эпидермиса на воздействие вращающихся
и переменных электрических полей низкой частоты// Гиг. и санит.- 1993.
- N 5. - С. 73-75.
12. Белкин А.Д. Устройство для исследования влияния вращающихся электрических
полей низкой частоты на лабораторных животных. А.с. N 2005407. Опубл. в
Б.И. - N 1. - 1994.
13. Белкин А.Д. Капсула для хранения веществ и культивирования
клеток. А.с. N 2012326. Опубл. в Б.И. - N 9 - 1994.
14. Белкин А.Д., Ясакова Н.Т., Тарабан В.Я., Мосолов А.Н. Влияние вращающихся
электрических полей промышленной частоты на систему крови лабораторных
животных// Гиг. и сан. - 1994. - N 9. - С. 32 - 33.
15. Белкин А.Д. Микобактерии в мембранной упаковке//Научная сессия
посвященная 100-летию Новосибирска, 25 - 28 января 1994 г.: Тез. докл.
Новосибирск, 1994. - С. 177.
16. Белкин А.Д. Один из возможных механизмов взаимодействия микобактерий
с фагоцитами/ 70 лет противотуберкулезной службе Новосибирской области.
50 лет Новосибирскому НИИ туберкулеза: сб. науч. тр. Юбилейной науч. -
практич. конф., Новосибирск, июнь 1995. - Новосибирск. - 1995.- С.
299 - 301.
17. Белкин А.Д. Эквивалентная электрическая схема человека с учетом
его анатомических особенностей//Актуальные вопросы современной медицины.
Тез. докл. Новосибирск, 1995.-Т.2.- С. 339-341.
18. Белкин А.Д., Бондарева А.А., Мосолов А.Н. Изменение микроядерного
теста и электропроводности под влиянием электрического поля промышленной
частоты у лабораторных животных// Гиг. и санит.- 1995.- N 2.- С. 43-44.
19. Белкин А.Д. Влияние вращающихся и переменных электрических
полей промышленной частоты на плодовитость, выживаемость и состояние внутренних
органов беспородных белых мышей// Гиг. и санит.- 1995.- N 4.- С.
28 - 32.
20. Белкин А.Д. О модульном принципе конструирования эквивалентных
электрических схем биоткани//2 съезд физиологов Сибири и Дальнего
Востока/Тез. докл. Новосибирск, 1995, часть 1. С. 43.
21. Белкин А.Д. Влияние вращающихся и переменных электрических полей
промышленной частоты на всхожесть овса//Тезисы докладов научной сессии
сотрудников НМИ. Новосибирск, 1995. С. 10.
22. Белкин А.Д. Влияние электрических полей на насекомых//Тезисы докладов
научной сессии сотрудников НМИ. Новосибирск, 1995. С. 11.
23. Белкин А.Д. О некоторых противоречиях в электромагнитной биологии//Тезисы
докладов научной сессии сотрудников НМИ. Новосибирск, 1995. С. 12.
24. Белкин А.Д. Влияние вращающихся и переменных электрических полей
на развитие трупного окоченения у мышей//Тезисы докладов научной сессии
сотрудников НМИ. Новосибирск, 1995. С. 15.
25. Белкин А.Д. Патофизиологические механизмы формирования эндоцитосимбиоза//Актуальные
вопросы патофизиологии лимфатической системы.- Новосибирск, 1995.- С. 74
- 77.
26. Белкин А.Д. О взаимодействии биосистем с электрическими полями
низкой частоты (теоретический аспект)//Гиг. и санит., 1996.- N 4.-
С. 27 - 31.
27. Белкин А.Д. Влияние вращающихся и переменных электрических
полей на течение острой циркуляторной гипоксии печени//Патогенез, профилактика
и коррекция гипоксических и ишемических состояний/Сб. ст. Под ред. А.В.Ефремова,
В.А.Шкурупия. - Новосибирск: Новосибирский медицинский институт, 1996.-
С. 7 - 8.
28. Белкин А.Д. О механизме канцерогенного действия электрических полей//Актуальные
вопросы современной медицины. Тез. докл. Новосибирск, 1996.-Т.2.- С. 260-261.
29. Ибрагимов Р.Ш., Белкин А.Д. О возможном механизме влияния электрических
полей низкой частоты на биологические объекты//Бюл. Сиб. отд-ния АМН СССР.
- 1989. - N 4. - С. 95 - 97.
30. Белкин А.Д. Способ воздействия на биологический объект. Патент
N 2076747. Опубл. в Бюл. N 10, 1997.
31. Белкин А.Д., Ибрагимов Р.Ш. Способ воздействия вращающимся электрическим
полем низкой частоты на лабораторных животных. Патент N 2076621. Опубл.
в Бюл. N 10, 1997.
32. Белкин А.Д. Новые звенья в патогенезе хронических инфекций//Врач.
1997.- N 7.- С. 40-41.
33. А.Д.Белкин. О роли вращающихся электрических полей в эндо- и экзоэкологических
взаимосвязях//3 съезд физиологов Сибири и Дальнего Востока/Тез.докл. Новосибирск,
1997. - С. 17.
